SISTEMA OCULOMOTORIO LENTO SMOOTH PURSUIT
Il sistema dei movimenti oculari di inseguimento lento (smooth pursuit) MOVIMENTI OCULARI
Nell’esame clinico del paziente la parte più importante riguarda l’esame dei movimenti oculari, ed in particolare la ricerca del nistagmo. fl nistagrno è definito come una serie di movimenti oculari (scosse) ciascuno costituito da una fase lenta che sposta il centro del bulbo verso la periferia, seguita da una fase rapida che riporta il bulbo al centro. La fase rapida denomina la direzione del nistagmo. Questo fenomeno se osservabile con capo fermo ed in posizione normale indica quasi sempre un disordine del sistema vestibolare e più precisamente un’alterazione del normale riflesso vestibolo-oculo motore (VOR).
Il VOR benché non richieda uno stimolo visivo, è tuttavia costantemente interattivo con gli stimoli visivi, e ciò ne aumenta notevolmente l’efficienza. Il VOR è necessario per stabilizzare la visione foveale durante i movimenti della testa. evitando così l’offuscamento delle immagini. I meccanismi centrali dell’oculomozione —sistema di inseguimento oculare lento, sistema saccadico. sistema otticocinetico. ed in misura minore il sistema di vergenza, combinano i loro effetti con il VOR per ottenere istante per istante la più nitida rappresentazione visiva del mondo circostante. Pertanto il VOR viene classificato entro la famiglia dei movimenti oculari che costantemente permettono di ottenere una visione distinta.
Nei paragrafi successivi si riporta una sintetica descrizione dei sistemi che presiedono ai vari tipi di movimento oculare classificati nella Tabella I,
CLASSIFICAZIONE MOVIMENTI OCULARI
|
Definizione |
Stimolo |
Funzione |
|
Inseguimento lento |
Bersagli lenti |
Fovea su bersagli visivi mobili |
|
Saccadici |
Bersagli rapidi |
|
|
Vergenza |
Bersagli frontali a distanza variabile |
|
|
VOR |
Piccoli movimenti del capo |
Fovea su bersagli mobili con capo in movimento |
|
Otticocinetici |
Successioni di bersagli o rotazione sostenuta ad occhi aperti |
|
Tab I. Cinque sistemi neuronali partecipano a mantenere la visione foveale Sistema di inseguimento lento mantiene l’immagine sula fovea mentre il bersaglio è in movimento. Sistema saccadico dirige rapidamente la fovea sul bersaglio che appare nella periferia del campo visivo. Sistema di vergenza muove i bulbi oculari in direzione opposta, cosicché la stessa immagine si proietta su entrambe le fovee (bersagli in movimento avanti-indietro). Sistema vestibolo-oculomotore dirige i bulbi oculari in direzione opposta a rotazioni brevi e veloci (2-4 Hz) della testa .Sistema ottico-cinetico’ usa informazioni visive per stabilizzare la fovea durante rotazioni della testa lente e prolungate
Test oculomotori: [ Il sistema dei movimenti oculari di inseguimento lento (smooth pursuit);il sistema saccadico ; movimenti di vergenza oculare; Il riflesso vestibolo oculo-motore; Il sistema otticocinetico ]
I movimenti d’inseguimento lento e saccadici possono essere facilmente studiati sotto videonistagmografia e/o videooculografia.. Essi permettono in pratica otorinolaringoiatrica di esplorare in pochi minuti in maniera non invasiva le vie oculogire e quindi le vie vestibolari centrali. Ultimamente si è assistito ad una rivalutazione dei movimenti oculari pursuit e saccadici nella pratica otoneurologica
anni 80 : lo studio dei movimenti oculari facevano parte integrante dell’esame vestibolare di II°
livello(ENG);per il loro contributo topo diagnostico e per il monitoraggio di malattie neurologiche e terapie associate Per provocare movimenti oculari coniugati di inseguimento, al
fine di studiare lo smooth pursuit, e saccadici possono essere utilizzate diverse tecniche di stimolazione che hanno in comune un input visivo caratterizzato da un bersaglio luminoso puntiforme in movimento lento su di un piano che coincide con quello dell’asse visivo diretto alternativamente verso un lato e l’altro con un movimento pendolare per il pursuit . La registrazione vera e propria dei saccadici si effettua facendo eseguire movimenti di fissazione di un led che si accende alternativamente con frequenza ed ampiezza random) sul piano orizzontale Il paziente, seduto ad una distanza di i metro dallo schermo, fissa il punto luminoso, in genere una “barra di LED”. La prova viene condotta in ambiente oscuro e silente. Il movimento oculare e lo spostamento del punto luminoso vengono separatamente registrati con ENG,
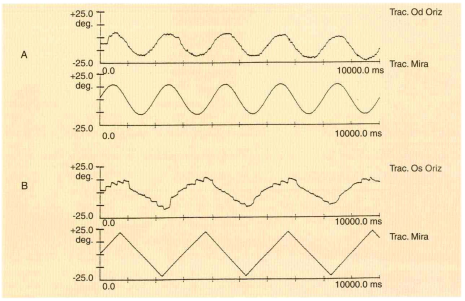
Fig. 1 – Movimento di inseguimento lento (tracciato superiore: movimento oculare; tracciato inferiore: movimento della mira).
A) movimenti di inseguimento normali; B) nello spostamento degli occhi verso destra, il movimento di inseguimento è sostituito da piccoli movimenti saccadici di rifissazione.

Fig. 2 – Vari tipi di anomalie dei movimenti saccadici (tracciato superiore: movimento oculare; tracciato inferiore: movimento della mira).
A) saccadi normali; B) dismetria saccadica (OD e OS tracciati superiori): i saccadici sono sostituiti da movimenti irregolari che raggiungono il target la cui fissazione non riesce ad essere mantenuta per tendenza dell’occhio a scivolare in posizione primaria. Microsaccadi di rifissazione per mantenere la fissazione del target o per correggere saccadi ipometriche; C) dismetria saccadica (poliradicoloneurite): ridotto spostamento dei bulbi oculari con tendenza allo scivolamento verso la posizione primaria di sguardo.
anni 90
Viene utilizzato lo stesso sistema di mire luminose o un proiettore LCD, soltanto che il sistema di registrazione ENG /VNG è dotato di apparecchiatura computerizzata per l’elaborazione delle tracce ottenute
|
|
|
|
|
|





anni 2000 Fig. 3a-b-c-d-e
Il test impulsivo cefalico (head impulse test – HIT)fig. 4 a-b-c-d fornisce una valutazione rapida e precisa, specifica per l’orecchio, del riflesso oculo-vestibolare (VOR) in risposta a stimoli nell’intervallo delle alte frequenze, l’intervallo naturale degli spostamenti del capo dandoci informazioni sui 3 canali semicircolari. fig. 5a-b


Un paziente con perdita vestibolare bilaterale
Test Impulsivo Cefalico ICS – Video Impulse test (vHIT)
|
|
|
|



paziente con ESAME DEI TRE CANALI SEMICIRCOLARI fig. 5a ICS

GRAFICO DEI 3 CANALI SEMICIRCOLARI D/S
Video Head Impulse Test (VHIT fig. 5b Snapsys)
Il sistema dei movimenti oculari di inseguimento lento (smooth pursuit)
Sistema di inseguimento lento o smooth pursuit
Anatomo-fisiologia
Lo smooth pursuit (SP) è una deviazione angolare dei bulbi oculari coniugata e riflessa che consente di mantenere sulla fovea un’immagine- bersaglio in movimento lento e continuo. elaborata dal sistema extra-genicolo-striato, per permettere al sistema genicolo striato l’osservazione foveale dell’immagine stessa. La funzione di tale riflesso è di fermare sulla fovea un’immagine-bersaglio in movimento lento.
Per fare questo, per mantenere sulla fovea l’immagine di un bersaglio che si muove lentamente nel campo visivo, lo Smooth Pursuit deve essere in grado di accoppiare la velocità degli occhi nello spazio (cioè la velocità dello sguardo) con la velocità del bersaglio, ma il sistema di inseguimento lento è anche capace di:
1.compensare per i movimenti della testa e/o dell’intero corpo che si verificano durante l’inseguimento lento in modo che la fovea possa seguire il bersaglio in movimento nello spazio;
2.cancellare il VOR se il bersaglio si muove in modo solidale con l’osservatore per mantenere l’immagine del target sulla fovea;
3.consentire l’inseguimento del bersaglio anche quando questo si muove oltre che sul piano frontale anche in profondità;
4.capire in anticipo quale sarà il movimento del bersaglio per sopperire alla lunga latenza dei movimenti di inseguimento (paragonata alla breve latenza del riflesso vestibolo-oculomotore, VOR, che è di circa 10 msec, quella dello SP è piuttosto lunga, circa 100 msec per i movimenti sul piano frontale e 150 msec per quelli in profondità).
Tutto questo suggerisce che esiste una stretta relazione funzionale non solo tra il sistema visivo e quello oculomotore, ma anche tra il sistema di smooth pursuit (sul piano frontale e/o in profondità)e quello vestibolare.
Lo start del sistema è a livello delle cellule gangliari retiniche (Figura 1) che proiettano le loro terminazioni alla corteccia visiva primaria (corteccia striata) tramite il nucleo genicolato laterale. A questo livello le aree visive occipitali corticali prendono connessioni con le aree extra-striate, (area mediale temporale, MT, e area mediale temporale superiore, MST), localizzate a livello della giunzione temporo-parieto-occipitale; queste ultime proiettano alla corteccia frontale, e precisamente ai frontal eye fields (FEFs) e ai supplementary eye fields (SEFs). Dalle aree extra-striate origina un fascio a proiezione puntina che rei mina a livello dei Nuclei Dorso-Laterali e Dorso-Mediali del Ponte (DLPNi, DMPNi); questi nuclei si connettono a loro volta controlateralmente con il fiocculo ed il verme posteriore del cervelletto. L’ultimo relè dell’arco riflesso neuronale è rappresentato dalle strutture premotorie (nuclei vestibolari ed integratori dei movimenti oculari), prima di giungere ai nuclei oculomotori dopo aver nuovamente attraversato la linea mediana. Il controllo del sistema cli SP dipende dalla conoscenza visiva del movimento della mira e si esplica attraverso il confronto fra l’elaborazione visiva (lei movimento della mii a ed il movimento di inseguimento lento.
Il movimento di pursuit avviene infatti in via riflessa tramite un meccanismo di pre-programmazione dei suoi parametri: infatti, se una mira luminosa in movimento a velocita costante viene impressivamente a scomparire, l’occhio continua ad eseguire ancora un movimenti) di pursuit valido per circa 200—300 msec.

Fig. 6 – Circuito neurale del movimento di SP a direzione orizzontale DLPN: Nuclei Pontini Dorso-Laterali; DMPN: Nuclei Pontini Dorso-Mediali; F: Flocculo; FEF: Frontal Eye Field; FN: Nuclei del Fastigium; LGN: Nucleo Genicolato Laterale; M: linea Mediana; MS: area Temporale Supero-Mediale; MT: area Temporale Mediale; MVN: Nucleo Vestibolare Mediale; NPH: Nucleo Preposito dell’ipoglosso; OVA: Aree Visive Occipitali; PAN: Nuclei Peri-Abducenti; PPC: Corteccia Parietale Posteriore; V: Verme; VI: nucleo abducente; I e 2: prima e seconda decussazione del circuito o: neuroni eccitatori; o: neuroni inibitori; linea tratteggiata: vie neuronali ipotetiche ancora da confermare
Affinché possa avere inizio l’inseguimento di una mira in movimento in qualsiasi parte del campo visivo, il sistema di SP deve poter accedere istante per istante all’analisi del movimento stesso (direzione e velocità); tale informazione viene fornita dalla MT, che possiede una mappa retinotopica del campo visivo controlaterale. Quando poi il movimento di inseguimento è cominciato, l’immagine del bersaglio viene a trovarsi sulla fovea o in prossimità di questa; a questo punto il sistema di Smooth Pursuit SP usa l’informazione visiva proveniente dalle regioni para-foveali per mantenere il movimento di inseguimento; ciò avviene grazie alla stimolazione visiva indotta dallo scivolamento dell’immagine del bersaglio sulla fovea, dovuto ad una non perfetta corrispondenza tra la velocità della mira e quella dello sguardo.

Fig.7 -Traccia EOG di movimento di SP. In questo grafico il “salto” (step) della mira di 5° verso destra e la susseguente “rampa” (ramp) in allontanamento con velocità di 16°/sec sono indicati dalla linea punteggiata Il tempo è sull’asse delle ascisse, la posizione su quello delle ordinate L’occhio (linea continua) segue il salto con un movimento rapido o saccadico per correggere l’errore di posizione e quindi prosegue con un movimento di inseguimento lento che mantiene il bersaglio vicino alla fovea Dato che il guadagno dell’arco riflesso neuronale è pari a 088, la mira tenderà ad allontanarsi dalla zona foveale durante il movimento di inseguimento, sarà allora compito di un saccade compensatorio riportare il bersaglio in zona parafoveale.
Il comando sul motoneurone oculare è di tipo step-ramp (Fig.7), nel quale il generatore fornirebbe lo step (scarica neurale costante e definita nel tempo necessaria a compiere un movimento rapido per portare la mira in prossimità della fovea) e l’integratore darebbe la conversione in un ramp (scarica neurale a frequenza progressiva definita nel tempo che consente di seguire la mira con un movimento lento e di mantenerla in zona foveale). L’arco riflesso alla base del pursuit è un circuito neurale ad anello chiuso con un guadagno di circa 0,88; tale guadagno è superiore a quello del VOR e pertanto, in caso di interazione negativa fra i due sistemi, l’input vestibolare viene ad essere annullato.
Il Movimento di Inseguimento Lento volontario, che si osserva però soltanto in presenza di piccoli bersagli ,che si muovono con moto uniforme e relativamente lento nel campo visivo, permette di mantenere sulle fovee della retina le immagini degli oggetti che si spostano nel campo visivo, attraverso un arco riflesso visuo-oculomotore, che comprende giunzioni corticali occipito-parietali e connessioni troncocerebellari Permette di stabilizzare un oggetto in  movimento sulla retina in modo da poterne esplorare i dettagli. Permette di “inseguire” con lo sguardo oggetti che si muovono “lentamente”, a circa 100°/sec Questo sistema di controllo interviene principalmente quando il bersaglio si muove e l’osservatore rimane fermo, oppure con bersaglio fermo e testa che ruota, ma anche con entrambi, testa e bersaglio in movimento con il fine di mantenere l’immagine continuamente sulla fovea della retina . Questa attitudine del sistema motorio dipende dal fatto che il sistema nervoso è in grado di CALCOLARE la DIREZIONE e VELOCITÀ del movimento dell’immagine del bersaglio sulla retina. Queste operazioni sono controllate dalla corteccia occipitale. . Con la testa ferma il movimento oculare è molto preciso, ma è evidente che nelle condizioni di testa in movimento, con bersaglio fisso o mobile, è necessaria un’integrazione fra VOR e sistema di inseguimento lento. Con testa e bersaglio entrambi in movimento, il sistema di inseguimento lento in realtà tende a sopprimere gli effetti del VOR. I movimenti di inseguimento oculare tendono a diventare imprecisi con l’età, per l’effetto di farmaci del s.n,c.. per la presenza di difetti dell’acuità visiva, per lesioni del cervelletto e delle corteccia occipitale e parietale.
movimento sulla retina in modo da poterne esplorare i dettagli. Permette di “inseguire” con lo sguardo oggetti che si muovono “lentamente”, a circa 100°/sec Questo sistema di controllo interviene principalmente quando il bersaglio si muove e l’osservatore rimane fermo, oppure con bersaglio fermo e testa che ruota, ma anche con entrambi, testa e bersaglio in movimento con il fine di mantenere l’immagine continuamente sulla fovea della retina . Questa attitudine del sistema motorio dipende dal fatto che il sistema nervoso è in grado di CALCOLARE la DIREZIONE e VELOCITÀ del movimento dell’immagine del bersaglio sulla retina. Queste operazioni sono controllate dalla corteccia occipitale. . Con la testa ferma il movimento oculare è molto preciso, ma è evidente che nelle condizioni di testa in movimento, con bersaglio fisso o mobile, è necessaria un’integrazione fra VOR e sistema di inseguimento lento. Con testa e bersaglio entrambi in movimento, il sistema di inseguimento lento in realtà tende a sopprimere gli effetti del VOR. I movimenti di inseguimento oculare tendono a diventare imprecisi con l’età, per l’effetto di farmaci del s.n,c.. per la presenza di difetti dell’acuità visiva, per lesioni del cervelletto e delle corteccia occipitale e parietale.
I movimenti di inseguimento lento (smooth pursuit eye movements) hanno una latenza di 100-150 ms.(fig. 8sm)
Dipendono però dall’aspettativa, e possono essere programmati in anticipo rispetto al movimento del bersaglio se esso si muove in modo prevedibile.
Questo sistema può operare solo quando l’immagine del bersagli è presente sulla retina, quindi non agisce al buio (al contrario del sistema saccadico).
Una delle differenze principali fra le saccadi ed i movimenti di inseguimento lento è il fatto che durante il movimento saccadico il nostro sistema visivo non acquisisce informazioni (soppressione
saccadica).
• Latenza: 100‐130 msec
• Velocità fino a 100°/sec
• Gain ottimale: 15‐30°/sec
• Caratteristiche variano per attenzione, motivazione,
età, velocità ed accelerazione del target, “memoria”
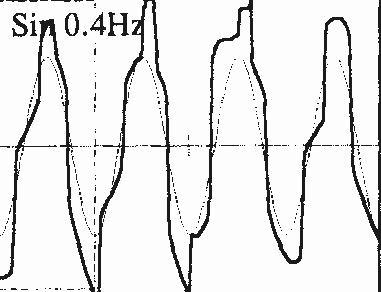
Il soggetto deve seguire un bersaglio che si sposta lentamente sul piano orizzontale da destra a sinistra e da sinistra a destra, oppure nel piano verticale dall’alto in basso e dal basso in alto. Lo stimolo ottimale in clinica è costituito da una stimolazione sinusoidale di 0,4 Hz, di ampiezza di più o meno 20° nel piano orizzontale e di 27° nel piano verticale. Il guadagno di questi movimenti di inseguimento (velocità oculare/velocità del bersaglio) è in seguito calcolato automaticamente e confrontato a delle norme prestabilite (cf. infra). Una riduzione del guadagno è un segno di interessamento centrale (Figura 8S.P.).

Aspetti clinici e strumentali delle alterazioni dello SP
Fig 8S.P. Movimenti oculari di inseguimento oculare lento anormali. La traccia nera rappresenta i movimenti del bersaglio. La traccia rossa rappresenta i movimenti oculari del paziente. Notare la riduzione del guadagno di questi movimenti in questo paziente.
Nella pratica clinica, il movimento oculare può risultare alterato o per patologia del sistema vestibolare che si ripercuote sul sistema di inseguimento lento (alterazione estrinseca) o per patologia diretta del sistema di inseguimento (alterazione intrinseca). Nella Tabella I sono riportati schematicamente i principali tipi di alterazione riscontrabili.
TABELLA II – Le diverse alterazioni dell’SP.
|
A) ALTERAZIONI ESTRINSECHE – Ny vestibolare • Ny spontaneo • Ny gaze evoked • Ny multiplo • Ny congenito |
B) ALTERAZIONI INTRINSECHE Guadagno ridotto Alterazioni in ampiezza |
Il nistagmo (Ny) vestibolare interferisce sul movimento oculare lento solo in due condizioni:
— quando il Ny non viene inibito dalla fissazione visiva (patologia vestibolare associata a patologia dell’arco riflesso di interazione visuo-vestibolare);
— quando il Ny ha un’ampiezza maggiore di 8°-10° o una velocità angolare maggiore di 20°-25°/s (incapacità dell’interazione visuo—vestibolare di sopprimere Ny vestibolari di tale intensità).
Quando la traccia sinusoidale viene compromessa da un Ny spontaneo, la sua morfologia si modifica in modo caratteristico (fig.9A). Per un Ny spontaneo sinistro, quando il movimento è da sinistra verso destra, l’occhio tende ad anticipare il bersaglio per un eccesso di guadagno neuronale, dovuto al fatto che il movimento di inseguimento è consensuale alla fase lenta del Ny, L’anticipo sul bersaglio viene corretto con un movimento saccadico di ritorno facilitato dalla fase rapida del Ny.
Quando il movimento di inseguimento è da destra verso sinistra, assistiamo ad un ritardo dell’occhio sul bersaglio legato al deficit di guadagno neuronale dovuto ad una interazione negativa con la fase lenta del Ny. L’occhio recupera l’esatta posizione sul bersaglio con un movimento saccadico di correzione facilitato dalla fase rapida del Ny.
Se è presente un Ny di lateralità bidirezionale, la traccia sinusoidale viene incisa ai due ventri dalla morfologia bifasica del Ny (Fig. 9B). La traccia fornisce inoltre una indicazione sulla simmetria o meno del Ny laterale. Ny di lateralità simmetrici si riscontrano frequentemente in lesioni centrali sistemiche di natura tossica o farmacologica, mentre un’asimmetria può essere espressione di lesione centrale emisisternica o prevalentemente emisistemica. In presenza di un Ny multidirezionale, il movimento oculare può divenire simil-atassico con una traccia EOG talvolta a morfologia quasi irriconoscibile (Fig.9C).

Fig. 9 – Registrazione EOG di SP a morfologia sinusoidale in presenza di Ny spontaneo sinistro (A), Ny di lateralità bilaterale (B), Ny multidirezionale (C), sostituzioni con movimenti saccadici multipli in condizioni di minore (D) o maggiore (E) grado di compromissione del sistema e in presenza di compromissione totale sia dell’SP che dei saccadici (F).
Particolare interesse neurologico rivestono le alterazioni del movimento di inseguimento lento per patologie dirette di questo sistema oculomotorio (alterazioni intrinseche). Quasi tutti i patterns EOG legati a patologia intrinseca del sistema sono caratterizzati da un ridotto guadagno neuronale dell’arco riflesso; l’alterazione di più frequente riscontro clinico è caratterizzata da sostituzioni della traccia sinusoidale di inseguimento con movimenti Saccadici multipli (Fig.9D); tale alterazione è espressione di un ridotto guadagno del sistema di inseguimento che determina un ritardo dell’occhio rispetto al movimento del bersaglio; il conseguente allontanamento dell’immagine dalla zona foveale attiva il sistema saccadico, determinando un movimento rapido compensatorio che riporta il bersaglio sulla fovea. I ripetuti saccadici di correzione che incidono la traccia sinusoidale sono la diretta conseguenza di una ridotta velocità angolare del movimento oculare lento, che non consente più un corretto inseguimento del bersaglio; non a caso la traccia EOG presenta il maggior numero di saccadici nella parte centrale dove è massima la velocità di spostamento del bersaglio e, se il guadagno neuronale del sistema di inseguimento è ulteriormente ridotto, l’intera parte centrale della traccia EOG viene sostituita da un unico saccadico (Figura 4E). L’alterazione della traccia sinusoidale per sostituzione ripetuta con saccadici, legata ad una disfunzione del sistema di inseguimento lento con relativa integrità del sistema saccadico, è espressione di sofferenza tronco-encefalica ed è comune a molti tipi di patologia (vascolare, flogistica, degenerativa, tumorale). Se esiste una compromissione sia del sistema di inseguimento che di quello saccadico, la traccia sinusoidale si presenta di ampiezza ridotta ed incostante, con saltuarie correzioni saccadiche. Questo tipo di pattern EOG si riscontra spesso nelle oftalmoplegie plus e nelle eredo-atassie (Fig.9F).
Nell’aprassia oculomotoria di Cogan si può ritrovare una completa assenza di movimenti saccadici con movimento di inseguimento lento parzialmente mantenuto; questo è l’unico caso in cui riscontriamo una traccia sinusoidale a morfologia normale, ma di ampiezza molto ridotta. L’aprassia oculomotoria di Cogan e la paralisi progressiva sopranucleare possono arrivare a determinare anche la completa scomparsa del movimento di inseguimento lento, che però è sempre preceduta da una più precoce scomparsa dei movimenti saccadici, prima sul piano verticale e poi su quello orizzontale; l’alterazione è in questi casi prevalentemente sopra mesencefalica, per lesione delle vie nervose occipito-mesencefaliche.
Disturbi simmetrici di Pursuit
Disturbi del Smooth Pursuit
L’età avanzata
Disturbi del tronco encefalico
Disturbi cerebellari
Disturbi cerebrali corticali
Nistagmo congenito
Ingestione droga
Disattenzione
Disturbi visivi
Pursuit asimmetrico
Cause di Pursuit asimmetrico
Malattia Acuta del lobo parietale
Malattia Acuta lobo frontale
Lesione dei nuclei pontini
Nistagmo sovrapposto
|
|
|
Fig. 10 Compromissione Simmetrica in un paziente con una lesione cerebellare ( impressione basilare ). |
Guadagno del Pursuit che è veramente superiore a 1,0 si verifica più frequentemente nei pazienti con una forma di nistagmo congenito chiamato “nistagmo latente”, durante la ricerca con onde triangolare. In Alcuni soggetti normali si possono registrare anche guadagni lievemente superiori a 1,0.
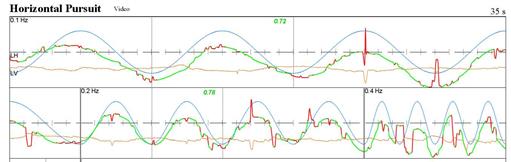
Bisogna avere una visione centrale per seguire lo stimolo luminoso. Così le persone non vedenti non possono seguire gli obiettivi, e le persone la cui visione è stata oscurata da una cataratta o da rimozione dei sistemi visivi di correzione. D’altra parte, pazienti con buona visione centrale, ma con cattiva visione periferica possono essere in grado di perseguire abbastanza bene. La figura seguente mostra un paziente che ha la visione periferica terribile (vedere la loro registrazione sotto la pagina di disturbo saccadici ), ma ha una buona visione centrale. Per questo motivo, possono perseguire abbastanza bene.
|
|
|
|
|
Fig. 11 I pazienti affetti da retinite pigmentosa (RP). In questa condizione la visione periferica è scarsa, ma la visione centrale può essere buono. Hanno “visione a tunnel”. Questo provoca una situazione particolare in cui saccadi possono essere molto disorganizzato, ma inseguimento può essere normale. |

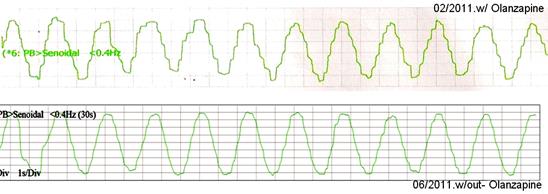
Viene riportato (Fig. 12) un esempio dell’effetto dell’ Olanzapina sul inseguimento (le immagini fornite da Dr. Dario Yacovino ).
DEL MOVIMENTO D’INSEGUIMENTO LENTO SMOOTH
Smooth pursuit Movimenti di inseguimento lento
Movimento di inseguimento lento che permette la continua foveazione di un oggetto che si muove lentamente nello spazio
• Latenza: 100‐130 msec
• Velocità fino a 100°/sec
• Gain ottimale: 15‐30°/sec
• Caratteristiche variano per attenzione, motivazione,
età, velocità ed accelerazione del target, “memoria”
Substrato anatomico dello smooth pursuit:
Retina ► tratto ottico accessorio ► sistema ottico accessorio ►
nuclei pretettali ► nucleo del tratto transpeduncolare (attività
spontanea che aumenta con il movimento lento del target)
Flocculo: riceve input vestibolari, visivi ed informazioni su
posizione e velocità oculare (gaze velocity Purkinje cells) ►
formazione reticolare pontina paramediana
Formazione reticolare (cancellazione del VOR, modulazione della
velocità)
Corteccia striata, parietale (area 7 ► input visivi transcorticali)
L’inseguimento oculare è un sistema di guida automatica dell’occhio su una mira in movimento. Si tratta di un adattamento molto preciso dei muscoli oculomotori per conservare una mira visiva sulla fovea retinica che si sposta lentamente nel campo visivo. L’inseguimento non è effettuato in ritardo rispetto alla mira ma in previsione; la velocità è infatti calcolata, stimata e corretta in base alla traiettoria dell’oggetto in movimento. Questo movimento inizia circa 200 millisecondi dopo la partenza della mira e persiste per 200 millisecondi dopo il suo arresto. Questo sistema si sviluppa relativamente tardi: fino alla sesta settimana di vita il bambino ha difficoltà a seguire con un movimento continuo l’oggetto che sta fissando, quando questo si muove. In genere compie una serie di movimenti saccadici.
Successivamente comincia a combinare movimenti rapidi e lenti con prevalenza sempre maggiore di quest’ultimi, fino a che dopo il 4° mese questo sistema risulta completamente sviluppato e permette di seguire oggetti in movimento (di tipo pendolare) fino ad una velocità di 30/40 °/sec, mentre ed oggetti che si muovono con un moto rettilineo fino a 100 °/sec.

Questo sistema di controllo interviene principalmente quando il bersaglio si muove e l’osservatore rimane fermo, con il fine di mantenere l’immagine continuamente sulla fovea. Questa attitudine del sistema motorio dipende dal fatto che il sistema nervoso è in grado di CALCOLARE la DIREZIONE e VELOCITÀ del movimento dell’immagine del bersaglio sulla retina. Queste operazioni sono controllate dalla corteccia occipitale.
Questo sistema può operare solo quando l’immagine del bersagli è presente sulla retina, quindi non agisce al buio (al contrario del sistema saccadico).
Nell’uomo, il movimento è uniforme e presenta un alto grado di regolarità se la velocità di spostamento dell’oggetto nel campo visivo è di circa 22°/sec. ma non superiore ai 30°/sec. Il sistema del pursuit in ambiente oscuro è inattivo.
Nello studio dei movimenti oculari di insegui mento i parametri numerici che dovranno essere considerati sono: la latenza (normale intorno a 130 msec.) ed il gain (velocità dell’occhio/ velocità del target). La velocità dell’occhio è in genere lievemente inferiore rispetto a quella del target, ma può essere influenzata da diversi fattori: attenzione, motivazione, età ,così come può essere influenzata da malattie o da farmaci.
In numerose patologie del sistema nervoso centrale i movimenti di inseguimento subiscono importanti alterazioni morfologiche o vengono sostituiti da irregolari movimenti saccadici di rifissazione del target (Fig. 13).
Numerosi sono i neuroni interessati nel meccanismo di controllo dei movimenti oculari. Si riscontrano nei nuclei vestibolari ,ma soprattutto nel tratto della sostanza reticolare paramediana del ponte compreso tra l’area nucleare vestibolare e i moto- neuroni oculari. Nella sostanza reticolare paramediana del ponte le unità fasiche che si attivano durante i movimenti saccadici sono prevalentemente localizzate nella parte rostrale, mentre quelle toniche che si attivano durante i movimenti lenti sono maggiormente presenti nella parte caudale attorno al nucleo dell’abducente. Durante la fissazione, tutti i saccadici vengono inibiti da cellule “pausa” disseminate ovunque nella sostanza paramediana pontina. La fissazione attiva le cellule “pausa” le quali, a loro volta, inibiscono le unità burst che determinano il saccadico.
Questo sistema di controllo interviene principalmente quando il bersaglio si muove e l’osservatore rimane fermo, con il fine di mantenere l’immagine continuamente sulla fovea.
I movimenti lenti o di inseguimento provocano il movimento degli occhi per mantenere sulla fovea l’immagine di un particolare bersaglio visivo che si muove nello spazio. Si tratta di movimenti volontari, per la cui esecuzione è necessaria la presenza di uno stimolo in movimento. Non è possibile eseguire un movimento lento di inseguimento in risposta ad un comando di natura esclusivamente verbale, senza la presenza di un bersaglio visivo in movimento; a differenza del nistagmo e dei movimenti saccadici, per effettuare questi movimenti è necessario prestare attenzione al bersaglio in movimento. La velocità massima è ritenuta essere di 100 °/sec., ma probabilmente questo valore è sovrastimato, ed uno più realistico potrebbe essere di 40 °/sec. per l’esecuzione dei movimenti.
Il test di controllo del sistema oculare per i movimenti di conduzione (pursuit,)
Nella routine otoneurologica, viene impiegato anche il test per l’analisi dei movimenti oculari lenti o di conduzione. Fin dalla prenotazione della visita, il paziente deve essere informato sulla durata dell’esame e sulle possibili interazioni farmacologiche con la funzione vestibolare. Bisogna inoltre raccomandargli di non assumere, 4 giorni prima, potenti farmaci con effetto sedativo vestibolare come la cinnarizina (Stugeron®), e la flunarizina (Sibelium®) con emivita di 18 giorni. Il Serc® ha un effetto sedativo vestibolare abbastanza esiguo (un’astensione di 24 ore appare più che sufficiente) mentre il Tanakan® e il Tanganil® non sono sedativi vestibolari. È più difficile astenersi dall’assumere neurolettici e sedativi; a volte, non è proprio possibile sospendere il Tegretol®, in caso di trattamento di una nevralgia facciale, e tantomeno un antiepilettico, farmaci che hanno effetti sulla funzione vestibolare e sull’oculomozione. Sempre a scopo cautelativo si ricorda che l’assunzione di alcol può dare piccoli segni di interessamento centrale; bisognerebbe dunque fare a meno di consumarne durante i pasti che precedono l’esame.
Vi sono due principali modalità di registrazione dei movimenti oculari: elettrica, con registrazione del potenziale corneoretinico: elettronistagmografia ed elettrooculografia; attraverso una telecamera a infrarossi che registra il movimento dell’occhio: videonistagmografia e suo complemento, video-oculografia.
L’occhio è un dipolo elettrico. Lo spostamento dell’occhio nell’orbita modifica il potenziale corneoretinico. Gli elettrodi posti sul piano orizzontale e sul piano verticale da una parte e dall’altra dell’occhio permettono di registrare questi spostamenti. La corrente generalmente captata, dell’ordine di 0,1 μV, viene amplificata. L’informatica ha permesso un’analisi precisa e affidabile della velocità delle fasi lente del nistagmo. L’elettronistagmografia permette di registrare le risposte oculari delle prove caloriche, delle prove rotatorie pendolari e del test di rotazione secondo un asse verticale inclinato rispetto alla gravità (RAIG), nonché i test oculografici (test di inseguimento oculare, test dei movimenti saccadici, test otticocinetici). Sul piano orizzontale e sul piano verticale di ciascun occhio vengono collocati degli elettrodi. La retina ha un’attività positiva rispetto alla cornea e il dipolo corneoretinico, il cui asse corrisponde più o meno a quello dell’asse ottico, fa variare dei campi elettrici perioculari durante questi spostamenti. Sulla pelle precedentemente decapata vengono collocati gli elettrodi in cloruro d’argento rivestiti di un sale d’argento. Una pasta abrasiva a base di granuli di pietra pomice permette di decapare la cute dopo averla sgrassata con miscela di alcol/etere; gli elettrodi vengono fissati con cerotti di piccole dimensioni.
L’elettronistagmografia e l’elettrooculografia erano in passato l’unico metodo in base al quale era possibile registrare i movimenti oculari. Queste tecniche sono attualmente sostituite, nell’adulto, dalla videonistagmografia e dalla videooculografia. Con la testa libera in ogni movimento, permettono una registrazione video dei movimenti oculari e un’analisi informatica delle risposte.
L’esame videonistagmografico è eseguito al buio con una telecamera sensibile agli infrarossi, miniaturizzata e disposta su una maschera completamente occlusiva. Questo esame permette di registrare i movimenti oculari spontanei, dopo le prove caloriche e rotatorie.
Nei test oculografici, è preferibile usare due telecamere in campo libero (una per ciascun occhio) con la possibilità di esaminare ogni occhio, mentre la testa del paziente è immobilizzata su una mentoniera. La video-oculografia consente la registrazione dei test di movimenti saccadici aleatori a 10°, 20° e 30° al secondo; in questo modo è possibile valutare eventuali ipo o ipermetrie, che fanno pensare a un’affezione cerebellare. Le prove dell’inseguimento oculare eseguite a 20°, 30° e 40° al secondo valutano la regolarità dei movimenti oculari. L’eventuale presenza di movimenti saccadici nell’inseguimento può segnalare anche in questo caso una lesione cerebellare.
Lo standard universale di queste telecamere è di 25 immagini al secondo; tuttavia, con un artificio di scansione dell’immagine, è possibile campionare fino a 50 emi-immagini al secondo servendosi separatamente di retini pari e dispari (procedimento Mumédia .

Per poter registrare questi movimenti oculari, non vi devono essere ammiccamenti, che andrebbero a interrompere il segnale o a modificare dei segnali elettrici. Può essere utile somministrare una goccia di Novesina® nella palpebra inferiore, per anestetizzare leggermente il paziente durante l’esame.
Le misure dei movimenti oculari richiedono una taratura. Il paziente deve guardare alternativamente delle mire con distanziamento ben controllato. Si tratta di una misurazione trigonometrica, dove la distanza del paziente rispetto alla mira è fondamentale. Lo spostamento oculare è una rotazione del globo; l’estrapolazione non è completamente lineare a partire da 30°-40° di disassiamento. Una calibratura più sfumata permette di correggere queste deformazioni.
Per convenzione, le deflessioni dei movimenti oculari verso destra sono rappresentate sulla carta o sullo schermo di visualizzazione verso l’alto; quelli verso sinistra saranno dunque diretti verso il basso; per quanto riguarda i movimenti verticali, la convenzione segue la logica del movimento, in alto verso l’alto, in basso verso il basso.
Il paziente, seduto ad una distanza di 1 metro dallo schermo o dalla mira luminosa fissa il punto luminoso. La velocità di spostamento del punto luminoso è di 5°/sec. La prova viene condotta in ambiente oscuro e silente. Il movimento oculare e lo spostamento del punto luminoso vengono registrati separatamente. In genere il gain non subisce alcuna alterazione in rapporto a variazioni della velocità di spostamento del segnale luminoso. Solo se viene superata la velocità (30°-60°/sec.) compatibile col sistema di controllo dei movimenti di conduzione, il tracciato apparirà interrotto per la presenza di saccadici.

Nel soggetto normale, anche i movimenti oculari di conduzione subiscono delle modificazioni a causa di variazioni fisiologiche del livello di attività della reticolare (stato di allerta o di riposo psicosensoriale, ecc.), di stati tossici (alcool) o dell’effetto di alcuni farmaci sedativi centrali. Anche l’età interferisce sulla regolarità dei movimenti lenti di inseguimento. Nei giovani (tra i 6 e 16 anni) e nelle persone anziane (tra 50 e 80 anni) è frequente osservare delle anomalie del pursuit mentre nei soggetti di media età (tra i 18 e 49 anni) i pursuit appaiono più accurati (Isotalo E., 1998). Il risultato normale è costituito da un movimento di inseguimento morbido, regolare e senza saccadi. Il principale risultato patologico è invece un inseguimento visivo saccadico a «scalini», riscontrato soprattutto nella patologia cerebellare. L’inseguimento può essere saccadico in una sola direzione (entrambi gli occhi); in tal caso si è in genere in presenza di una lesione cerebellare omolaterale .
Nella patologia del SNC, le alterazioni del pursuit si osservano nelle lesioni emisferiche, dei gangli della base e in quelle del cervelletto. Se il danno è diffuso, le anomalie dei movimenti di conduzione si osservano bilateralmente. Se invece la lesione è unilaterale (a livello del lobo parieto-occipitale o di un emisfero cerebellare oppure del tronco encefalico) o è limitata alla fossa cranica posteriore di un lato, l’alterazione del pursuit è omolaterale alla sede del focolaio. Per l’anomalia del movimento oculare di conduzione, il paziente è spesso costretto ad eseguire dei piccoli movimenti degli occhi che si rendono necessari per mantenere il punto luminoso sulla fovea. Ciò si verifica con valori di spostamento del punto luminoso di bassa velocità (10°/sec.). In questi casi, la curva sinusoide del pursuit assume un aspetto seghettato per l’interferenza dei saccadici (alterazione “a scala” di Maspetiol). Altre volte, la curva sinusoide del pursuit appare irregolare per la sovrapposizione di un nistagmo da “direzione dello sguardo” allorché gli occhi ruotano verso il lato dove questo è presente. Tale irregolarità del pursuit può essere presente in ambedue le estremità della curva sinusoide se il nistagmo da “direzione dello sguardo” è bilaterale. Analogo aspetto può assumere il pursuit qualora venga evocato sul piano verticale in presenza di un nistagmo da “direzione dello sguardo” quando è diretto verso l’alto più raramente verso il basso.
In caso di oftalmoplegia internucleare associata a una patologia cerebellare, l’inseguimento è saccadico solo nell’occhio diretto in abduzione.
In caso di nistagmo congenito, l’inseguimento diventa patognomonico e associa una successione di movimenti oculari non saccadici, ma pseudociclici (alla registrazione elettronistagmografica si ha un aspetto a cupola e a pagoda).
L’ultima prova che conferma la patologia sopranucleare è l’assenza di anomalie nella manovra degli «occhi di bambola». Quest’esame consiste nel far oscillare passivamente la testa del paziente nel piano orizzontale, invitandolo a osservare un punto fisso davanti a sé. Tali movimenti non costituiscono più un inseguimento visivo ma un adattamento vestibolare. Questa è la prova in base alla quale si può affermare che la via comune dell’oculomozione è indenne.
Gli errori di valutazione nell’inseguimento saccadico sono principalmente causati dalle seguenti situazioni:
■ il paziente è subvedente;
■ presenta un nistagmo congenito insolito;
■ presenta strabismo e cambia l’occhio che fissa durante l’inseguimento Fig. 16
|
|
|
Fig. |


Bibliografia
J. Fukushima, T. Akao, S. Kurkin, C.R.S. Kaneko, K. Fukushima. The vestibular-related frontal cortex and its role in smooth pursuit eye movements and vestibular-pursuit interactions. J Vestib Res, 2006; 16(1-2): 1-22.
M. Versino, S. Colnaghi. Il saccadico nelle malattie del SNC. Atti congressuali “Aggiornamenti di vestibologia”,3a Edizione; Modena, 10-11 Marzo 2006.
A.F. Fuchs, F.R. Robinson, A Straube. Role of the Caudal Fastigial Nucleus i Saccade Generation. I.Neuronal Discharge Patterns Journal of Neurophysiology, 1993; 70(5): 1723-1740.
S. Ramat, R.J. Leigh, D.S. Zee. L.M. Optican. What clinical disorders tell us about the neural control of saccadic eye movements. Brain, 2007; 130: 10-35.
S. Traccis, D. Zambarbieri. I movimenti di inseguimento lento. Pàtron Editore, Bologna 1994.
S. Traccis, D. Zambarbieri. I movimenti saccadici. Pàtron Editore, Bologna 1992.
R.J. Leigh, D.S. Zee. The neurology of eye movements. 2nd ed., F.A. Davis Company, Philadelphia 1991.
U. Büttner, T. Brandt. Ocular motor disorders of the brain stem. Vol. 1, Nr. 2, August 1992, “Clinical Neurology”, Baillière Tindall, London.
P. Pagnini. Interazioni visuo-vestibolo-oculomotorie. da “Manuale di Audiologia” di M. Del Bo, F. Giaccai,
G. Grisanti, 2a ed., Masson Italia Editori, Milano 1994.
P. Pagnini. Patterns EOG visuo-oculomotori nella patologia tronco-cerebellare. Atti del Simposio
Internazionale “Potenziali evocati uditivi nella patologia del sistema nervoso centrale”, C.R.S. Amplifon, Milano, 26-27 Novembre 1982.
F. Giaccai, P. Pagnini. Patterns elettronistagmografici nelle sindromi vestibolari centrali. da “Les
explorations functionelles objectives en O.R.L.”, relazione al XXIII Congresso della Società Latina di O.R.L., Bruxelles 1980.TrattatodiVestibologia
BIBLIOGRAFIA
- Adler SA, Bala J & Krauzlis RJ (2002) Primato delle informazioni spaziali nel guidare la selezione del target per l’esercizio e la saccadi J Vision 2, 627 -.. 644.
- Bahill AT, Clark MR & Stark L (1975) La sequenza principale, uno strumento per lo studio dei movimenti oculari umani Math Biosci 24, 191 -.. 204.
- Bahill AT & McDonald JD (1983) Smooth movimenti perseguimento degli occhi in risposta alle prevedibili movimenti bersaglio Vision Res 23, 1573 -.. 1583.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Barnes & GR Asselman PT (1991) Il meccanismo di previsione dei movimenti oculari ricerca lisce umane J Physiol 439, 439 -.. 461.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Barnes G, Goodbody S & S Collins (1995) controllo volontario delle risposte anticipatorie inseguimento oculari in condizioni stabilizzate immagine nell’uomo Exp Brain Res 106, 301 -.. 317.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Basso MA, Krauzlis RJ & Wurtz RH (2000) Attivazione e l’inattivazione di rostrali neuroni collicolo superiore durante movimenti oculari liscio-inseguimento nelle scimmie J Neurophysiol 84, 892 -.. 908.
- PubMed ,
- CAS ,
- Web of Science®
- . Basso MA, Pokorny JJ Liu & P (2005) Attività della sostanza nera pars reticulata neuroni durante i movimenti oculari ricerca lisci nelle scimmie Eur J Neurosci 22, 448 -. 464.
Link Diretto:
- Becker W (1991). Saccades. .. In Eye Movements, ndr CarpenterR, pp 95-137. MacMillan Press, Houndmills, Regno Unito.
- Becker W & Fuchs AF (1985) Previsione del sistema oculomotore: esercizio regolare durante scomparsa transitoria di un bersaglio visivo Exp Brain Res 57, 562-575…
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Bennett SJ & Barnes GR (2003) inseguimento oculare umano durante la scomparsa transitoria di un bersaglio visivo J Neurophysiol 90, 2504 -.. 2520.
- Bennett SJ, Orban de Xivry JJ, Barnes GR & Lefevre P (2007) Accelerazione bersaglio può essere estratto e rappresentato all’interno dell’unità predittivo per oculare ricerca J Neurophysiol; 98 (3), 1405-1414…
- Berman RA, Colby CL, Genovese CR, Voyvodic JT, Luna B, Thulborn KR & Sweeney JA (1999) le reti corticali che sottendono inseguimento e movimenti oculari saccadici nell’uomo: uno studio fMRI Hum Cervello Mapp 8, 209-225…
Link Diretto:
- Blohm G, Messale M & P Lefevre (2003a) Interazione tra anticipazione liscia e saccadi durante l’orientamento oculare nelle tenebre J Neurophysiol 89, 1423 -.. 1433.
- . Blohm G, Messale M & P Lefevre (2003b) Smooth movimenti oculari anticipatori alterano la posizione memorizzata di obiettivi balenavano J Vis 3, 761 -. 770.
- Blohm G, Messale M & P Lefevre (2005a) Elaborazione dei segnali retinici e extraretinal per saccadi memoria guidato durante l’esercizio regolare J Neurophysiol 93, 1510 -.. 1522.
- Blohm G, Messale M & P Lefevre (2005b) la prova diretta per un ingresso di posizione al sistema di ricerca liscio J Neurophysiol 94, 712 -.. 721.
- . Blohm G, Optican LM & P Lefevre (2006) Un modello che integra la velocità occhio comandi per tenere traccia degli spostamenti degli occhi liscio J Comput Neurosci 21, 51 -. 70.
- Boeddeker N & Egelhaaf M (2005) un sistema di controllo unico per lo svolgimento regolare e saccade-come in mosconi J Exp Biol 208, 1563 -.. 1572.
- Bogousslavsky J & Meienberg O (1987) i disturbi dei movimenti oculari in tronco cerebrale e ictus cerebellare Arch Neurol 44, 141 -.. 148.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Nato RT & Bradley DC (2005) Struttura e funzione di area visiva MT Annu Rev Neurosci 28, 157 -.. 189.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Braun DI, Pracejus L & Gegenfurtner KR (2006) Proposta collaterale suscita movimenti oculari inseguimento liscio J Vis 6, 671 -.. 684.
- Bridgeman B (1995) Una revisione del ruolo della copia efferenza nei sistemi di controllo sensoriali e oculomotorie Ann Biomed Eng 23, 409 -.. 422.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Bridgeman B, Hendry D & L Stark (1975) Mancata rivelare lo spostamento del mondo visivo durante i movimenti oculari saccadici Vision Res 15, 719 -.. 722.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- . Carello CD & Krauzlis RJ (2004) Manipolazione intento: prova per un ruolo causale del collicolo superiore nella scelta di destinazione Neuron 43, 575-583..
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Carl JR & Gellman RS (1987) umana inseguimento liscio: risposte stimolo-dipendente J Neurophysiol 57, 1446-1463…
- PubMed ,
- CAS ,
- Web of Science®
- Carpenter RH & Williams ML (1995) il calcolo neurale di log probabilità nel controllo dei movimenti oculari saccadici [vedere Commenti] Nature 377, 59 -.. 62.
- CrossRef ,
- PubMed ,
- Web of Science® ,
- ADS
- Chubb MC & Fuchs AF (1982) Contributo di y gruppo di nuclei vestibolari e nucleo dentato del cervelletto di generazione di movimenti oculari verticali lisce J Neurophysiol 48, 75 -.. 99.
- PubMed ,
- CAS ,
- Web of Science®
- Churchland AK & Lisberger SG (2002) Controllo di guadagno in movimenti oculari liscia inseguimento umani J Neurophysiol 87, 2936 -.. 2945.
- Churchland MM, Chou IH & Lisberger SG (2003) Prove per oggetto la permanenza nei movimenti oculari liscia perseguimento di scimmie J Neurophysiol 90, 2205 -.. 2218.
- Cui DM, Yan YJ & Lynch JC (2003) Pursuit subregione dei progetti frontali campo occhio al nucleo caudato nelle scimmie J Neurophysiol 89, 2678 -.. 2684.
- Dallos PJ & Jones RW (1963) Apprendimento comportamento del sistema di controllo fissazione dell’occhio IEEE Trans controllo automatico 8, 218 -.. 227.
- De Brouwer S, M Messale, Barnes G & P Lefevre (2002a) Analisi quantitativa di saccadi catch-up durante l’esercizio prolungato J Neurophysiol 87, 1772 -.. 1780.
- De Brouwer S, Messale M & P Lefevre (2001) Il ruolo di slittamento della retina nella previsione del movimento di destinazione durante l’esercizio regolare e saccadico J Neurophysiol 86, 550 -.. 558.
- PubMed ,
- CAS ,
- Web of Science®
- De Brouwer S, Yuksel D, Blohm G, Messale M & P Lefevre (2002b) Che cosa fa scattare saccadi catch-up durante il monitoraggio visivo J Neurophysiol 87, 1646 -.? 1650.
- Dicke PW, Barash S, Ilg UJ & Thier P (2004) la prova singolo neurone per un contributo dei nuclei pontini dorsale di entrambi i tipi di movimenti oculari bersaglio diretto, saccadi e liscia esercizio Eur J Neurosci 19, 609.. – 624.
Link Diretto:
- . Dürsteler MR & Wurtz RH (1988) Pursuit e deficit optokinetic a seguito di lesioni chimiche delle aree corticali MT e MST J Neurophysiol 60, 940 -. 965.
- PubMed ,
- CAS ,
- Web of Science®
- Erkelens CJ (2006) Coordinamento della ricerca liscia e saccadi Vision Res 46, 163 -.. 170.
- Fischer B & Boch R (1983) i movimenti oculari saccadici dopo tempi di reazione estremamente brevi scimmia Brain Res 260, 21 -.. 26.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Fuchs AF, Robinson FR & Straube A (1993). Il ruolo del nucleo fastigial caudale in generazione saccade. I. neuronale modello scarico J Neurophysiol 70, 1723 -. 1740.
- PubMed ,
- CAS ,
- Web of Science®
- Fuchs AF, Robinson FR & Straube A (1994). La partecipazione del nucleo fastigial caudale in movimenti oculari liscia inseguimento. . I. neuronale attività J Neurophysiol 72, 2714-2728.
- PubMed ,
- CAS ,
- Web of Science®
- Gardner JL & Lisberger SG (2001) selezione del target collegato ai movimenti oculari saccadici ricerca e lisce J Neurosci 21, 2075 -.. 2084.
- PubMed ,
- CAS ,
- Web of Science®
- Gellman RS & Carl JR (1991) il trattamento Proposta di movimenti oculari saccadici nell’uomo Exp Brain Res 84, 660 -.. 667.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- .. Guan Y, Eggert T, Bayer O & Büttner U (2005) Saccades agli obiettivi fissi e in movimento differiscono nella scimmia Exp Brain Res 161, 220-232.
- Hanes DP & Carpenter RH (1999) contrordine saccades nell’uomo Vision Res 39, 2777 -.. 2791.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Hanes DP & Schall JD (1995) contrordine saccades in macaco Vis Neurosci 12, 929 -.. 937.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- . Harris CM & Wolpert DM (1998) di rumore del segnale in funzione determina pianificazione motoria Natura 394, 780 -. 784.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science® ,
- ADS
- . Harris CM & Wolpert DM (2006) La sequenza principale di saccadi ottimizza velocità precisione trade-off Biol Cybern 95, 21 -. 29.
- Harting JK (1977) vie discendenti dal collicullus superiore: analisi autoradiografica nelle scimmie rhesus (Macaca mulatta) J Neurol Comp 173, 583-612…
Link Diretto:
- Heinen SJ & Liu M (1997) l’attività Single-neuroni nella corteccia frontale dorsomediale durante i movimenti oculari liscio-inseguimento per prevedibile movimento obiettivo Vis Neurosci 14, 853 -.. 865.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Ilg UJ & Churan J (2004) la percezione di movimento senza attività esplicito nelle aree MT e MST J Neurophysiol 92, 1512 -.. 1523.
- Joiner WM & Shelhamer M (2006) Pursuit e saccadico mostra inseguimento una dipendenza simile tempo di preparazione movimento Exp Brain Res 173, 572 -.. 586.
- Keller E & Johnsen SD (1990) la previsione di velocità in saccadi correttive durante i movimenti oculari liscio-inseguimento in scimmia Exp Brain Res 80, 525 -.. 531.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Keller EL, Gandhi NJ & Shieh JM (1996a) accuratezza Endpoint in saccades interrotte da stimolazione nella regione omnipause in scimmia Vis Neurosci 13, 1059 -.. 1067.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Keller EL, Gandhi NJ & Weir PT (1996b) Scarico di neuroni collicular superiori durante le saccadi apportate a bersagli in movimento J Neurophysiol 76, 3573 -.. 3577.
- PubMed ,
- CAS ,
- Web of Science®
- Keller EL & Messale M (2003) percorsi del tronco cerebrale condivise per saccades e movimenti oculari liscia inseguimento Ann NY Acad Sci 1004, 29 – 39…
Link Diretto:
- Kim CE, Thaker GK, Ross DE & Medoff D (1997) precisioni di saccadi a bersagli in movimento durante l’esercizio e la manutenzione di iniziazione Exp Brain Res 113, 371 -.. 377.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Knox PC (1996) L’effetto del paradigma gap sulla latenza del liscio ricerca umana del movimento oculare Neuroreport 7, 3027 -.. 3030.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Komatsu H & Wurtz RH (1989) di modulazione dei movimenti oculari ricerca attraverso la stimolazione delle aree corticali MT e MST J Neurophysiol 62, 31 -.. 47.
- PubMed ,
- CAS ,
- Web of Science®
- Kornylo K, Dill N, Saenz M & Krauzlis RJ (2003) Annullamento di movimenti oculari saccadici e pursuit negli esseri umani e scimmie J Neurophysiol 89, 2984 -.. 2999.
- Krauzlis RJ (2003) l’attività neuronale in collicolo rostrale superiore relative all’avvio di ricerca e movimenti oculari saccadici J Neurosci 23, 4333 -.. 4344.
- PubMed ,
- CAS ,
- Web of Science®
- .. Krauzlis RJ (2004) di revisione del sistema perseguimento movimento oculare liscio J Neurophysiol 91, 591-603.
- Krauzlis RJ (2005) Il controllo dei movimenti oculari volontari: nuove prospettive neuroscienziato 11, 124-137…
- Krauzlis RJ, Basso MA & Wurtz RH (1997) Errore motore comune per molteplici movimenti oculari Science 276, 1693 -.. 1695.
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Krauzlis R & Dill N ( 2002 ). Neural correlates of target choice for pursuit and saccades in the primate superior colliculus . Neuron 35 , 355 – 363 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Krauzlis RJ & Lisberger SG ( 1994 ). A model of visually-guided smooth pursuit eye movements based on behavioral observations . J Comput Neurosci 1 , 265 – 283 .
- Krauzlis RJ & Miles FA ( 1996a ). Decreases in the latency of smooth pursuit and saccadic eye movements produced by the ‘gap paradigm’ in the monkey . Vision Res 36 , 1973 – 1985 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Krauzlis RJ & Miles FA ( 1996b ). Release of fixation for pursuit and saccades in humans: evidence for shared inputs acting on different neural substrates . J Neurophysiol 76 , 2822 – 2833 .
- PubMed ,
- CAS ,
- Web of Science®
- Krauzlis RJ & Miles FA ( 1998 ). Role of the oculomotor vermis in generating pursuit and saccades: effects of microstimulation . J Neurophysiol 80 , 2046 – 2062 .
- PubMed ,
- CAS ,
- Web of Science®
- Krauzlis RL , Zivotofsky AZ & Miles FA ( 1999 ). Target selection for pursuit and saccadic eye movements in humans . J Cogn Neurosci 11 , 641 – 649 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Kurylo DD & Skavenski AA ( 1991 ). Eye movements elicited by electrical stimulation of area PG in the monkey . J Neurophysiol 65 , 1243 – 1253 .
- PubMed ,
- CAS ,
- Web of Science®
- Lefevre P & Galiana HL ( 1992 ). Dynamic feedback to the superior colliculus in a neural network model of the gaze control system . Neural Networks 5 , 871 – 890 .
- Lefevre P , Missal M & Galiana HL ( 1994 ). Modeling slow correcting gaze movements . J Vestib Res 4 , 371 – 381 .
- Leigh RJ & Kennard C ( 2004 ). Using saccades as a research tool in the clinical neurosciences . Brain 127 , 460 – 477 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Leigh JR & Zee DS ( 2006 ). The Neurology of Eye Movements . Oxford University Press Inc., New York .
- Lisberger SG , Morris EJ & Tychsen L ( 1987 ). Visual motion processing and sensory-motor integration for smooth pursuit eye movements . Annu Rev Neurosci 10 , 97 – 129 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Lisberger SG & Movshon JA ( 1999 ). Visual motion analysis for pursuit eye movements in area MT of macaque monkeys . J Neurosci 19 , 2224 – 2246 .
- PubMed ,
- CAS ,
- Web of Science®
- Lisberger SG & Westbrook LE ( 1985 ). Properties of visual inputs that initiate horizontal smooth pursuit eye movements in monkeys . J Neurosci 5 , 1662 – 1673 .
- PubMed ,
- CAS ,
- Web of Science®
- Liston D & Krauzlis RJ ( 2003 ). Shared response preparation for pursuit and saccadic eye movements . J Neurosci 23 , 11305 – 11314 .
- PubMed ,
- CAS ,
- Web of Science®
- Liston D & Krauzlis RJ ( 2005 ). Shared decision signal explains performance and timing of pursuit and saccadic eye movements . J Vis 5 , 678 – 689 .
- Logan GD , Cowan WB & Davis KA ( 1984 ). On the ability to inhibit simple and choice reaction time responses: a model and a method . J Exp Psychol Hum Percept Perform 10 , 276 – 291 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Madelain L & Krauzlis RJ ( 2003 ). Effects of learning on smooth pursuit during transient disappearance of a visual target . J Neurophysiol 90 , 972 – 982 .
- Masson GS & Stone LS (2002) Dal seguente bordi per gli oggetti che perseguono J Neurophysiol 88, 2869 -.. 2873.
- May JG , Keller EL & Suzuki DA ( 1988 ). Smooth-pursuit eye movement deficits with chemical lesions in the dorsolateral pontine nucleus of the monkey . J Neurophysiol 59 , 952 – 977 .
- PubMed ,
- CAS ,
- Web of Science®
- Merrison AF & Carpenter RH ( 1995 ). ‘Express’ smooth pursuit . Vision Res 35 , 1459 – 1462 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Missal M , Coimbra A , Lefevre P & Olivier E ( 2002a ). A quantitative analysis of the correlations between eye movements and neural activity in the pretectum . Exp Brain Res 143 , 373 – 382 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Missal M , Coimbra A , Lefevre P & Olivier E ( 2002b ). Further evidence that a shared efferent collicular pathway drives separate circuits for smooth eye movements and saccades . Exp Brain Res 147 , 344 – 352 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Missal M , Crommelinck M , Roucoux A & Decostre MF ( 1993 ). Slow correcting eye movements of head-fixed, trained cats toward stationary targets . Exp Brain Res 96 , 65 – 76 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Missal M , De Brouwer S , Lefevre P & Olivier E ( 2000 ). Activity of mesencephalic vertical burst neurons during saccades and smooth pursuit . J Neurophysiol 83 , 2080 – 2092 .
- PubMed ,
- CAS ,
- Web of Science®
- Missal M & Keller EL ( 2002 ). Common inhibitory mechanism for saccades and smooth-pursuit eye movements . J Neurophysiol 88 , 1880 – 1892 .
- PubMed ,
- CAS ,
- Web of Science®
- Missal M , Lefevre P , Crommelinck M & Roucoux A ( 1995 ). Evidence for high-velocity smooth pursuit in the trained cat . Exp Brain Res 106 , 509 – 512 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Missal M , Lefevre P , Delinte A , Crommelinck M & Roucoux A ( 1996 ). Smooth eye movements evoked by electrical stimulation of the cat’s superior colliculus . Exp Brain Res 107 , 382 – 390 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Mitrani L & Dimitrov G ( 1978 ). Pursuit eye movements of a disappearing moving target . Vision Res 18 , 537 – 539 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Morris EJ & Lisberger SG ( 1987 ). Different responses to small visual errors during initiation and maintenance of smooth-pursuit eye movements in monkeys . J Neurophysiol 58 , 1351 – 1369 .
- PubMed ,
- CAS ,
- Web of Science®
- Newsome WT , Wurtz RH , Dursteler MR & Mikami A ( 1985 ). Deficits in visual motion processing following ibotenic acid lesions of the middle temporal visual area of the macaque monkey . J Neurosci 5 , 825 – 840 .
- PubMed ,
- CAS ,
- Web of Science®
- Nijhawan R ( 1994 ). Motion extrapolation in catching . Nature 370 , 256 – 257 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science® ,
- ADS
- Orban de Xivry JJ , Bennett SJ , Lefevre P & Barnes GR ( 2006 ). Evidence for synergy between saccades and smooth pursuit during transient target disappearance . J Neurophysiol 95 , 418 – 427 .
- Osman A , Kornblum S & Meyer DE ( 1986 ). The point of no return in choice reaction time: controlled and ballistic stages of response preparation . J Exp Psychol Hum Percept Perform 12 , 243 – 258 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Petit L & Haxby JV ( 1999 ). Functional anatomy of pursuit eye movements in humans as revealed by fMRI . J Neurophysiol 82 , 463 – 471 .
- PubMed ,
- CAS ,
- Web of Science®
- Price NS , Ono S , Mustari MJ & Ibbotson MR ( 2005 ). Comparing acceleration and speed tuning in macaque MT: physiology and modeling . J Neurophysiol 94 , 3451 – 3464 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Priebe NJ , Churchland MM & Lisberger SG ( 2001 ). Reconstruction of target speed for the guidance of pursuit eye movements . J Neurosci 21 , 3196 – 3206 .
- PubMed ,
- CAS ,
- Web of Science®
- Priebe NJ & Lisberger SG ( 2004 ). Estimating target speed from the population response in visual area MT . J Neurosci 24 , 1907 – 1916 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Ramat S , Leigh RJ , Zee DS & Optican LM ( 2007 ). What clinical disorders tell us about the neural control of saccadic eye movements . Brain 130 , 10 – 35 .
- Rashbass C ( 1961 ). The relationship between saccadic and smooth tracking eye movements . J Physiol 159 , 326 – 338 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Robinson DA , Gordon JL & Gordon SE ( 1986 ). A model of the smooth pursuit eye movement system . Biol Cybern 55 , 43 – 57 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Rosano C , Krisky CM , Welling JS , Eddy WF , Luna B , Thulborn KR & Sweeney JA ( 2002 ). Pursuit and saccadic eye movement subregions in human frontal eye field: a high-resolution fMRI investigation . Cereb Cortex 12 , 107 – 115 .
- Schoppik D & Lisberger SG ( 2006 ). Saccades exert spatial control of motion processing for smooth pursuit eye movements . J Neurosci 26 , 7607 – 7618 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Schreiber C , Missal M & Lefevre P ( 2006 ). Asynchrony between position and motion signals in the saccadic system . J Neurophysiol 95 , 960 – 969 .
- Segraves MA & Goldberg ME ( 1994 ). Effect of stimulus position and velocity upon the maintenance of smooth pursuit eye velocity . Vision Res 34 , 2477 – 2482 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Sommer MA ( 2003 ). The role of the thalamus in motor control . Curr Opin Neurobiol 13 , 663 – 670 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Sommer MA & Wurtz RH ( 2002 ). A pathway in primate brain for internal monitoring of movements . Science 296 , 1480 – 1482 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science® ,
- ADS
- Sommer MA & Wurtz RH ( 2004a ). What the brain stem tells the frontal cortex. I. Oculomotor signals sent from superior colliculus to frontal eye field via mediodorsal thalamus . J Neurophysiol 91 , 1381 – 1402 .
- Sommer MA & Wurtz RH ( 2004b ). What the brain stem tells the frontal cortex. II. Role of the SC-MD-FEF pathway in corollary discharge . J Neurophysiol 91 , 1403 – 1423 .
- Steinbach MJ ( 1976 ). Pursuing the perceptual rather than the retinal stimulus . Vision Res 16 , 1371 – 1376 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Takagi M , Zee DS & Tamargo RJ ( 1998 ). Effects of lesions of the oculomotor vermis on eye movements in primate: saccades . J Neurophysiol 80 , 1911 – 1931 .
- PubMed ,
- CAS ,
- Web of Science®
- Takagi M , Zee DS & Tamargo RJ ( 2000 ). Effects of lesions of the oculomotor cerebellar vermis on eye movements in primate: smooth pursuit . J Neurophysiol 83 , 2047 – 2062 .
- PubMed ,
- CAS ,
- Web of Science®
- Tanaka M ( 2005 ). Involvement of the central thalamus in the control of smooth pursuit eye movements . J Neurosci 25 , 5866 – 5876 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Tanaka M & Lisberger SG ( 2001 ). Regulation of the gain of visually guided smooth-pursuit eye movements by frontal cortex . Nature 409 , 191 – 194 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science® ,
- ADS
- Thier P & Ilg UJ ( 2005 ). The neural basis of smooth-pursuit eye movements . Curr Opin Neurobiol 15 , 645 – 652 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Tian JR & Lynch JC ( 1996 ). Functionally defined smooth and saccadic eye movement subregions in the frontal eye field of Cebus monkeys . J Neurophysiol 76 , 2740 – 2753 .
- PubMed ,
- CAS ,
- Web of Science®
- Todorov E ( 2004 ). Optimality principles in sensorimotor control . Nat Neurosci 7 , 907 – 915 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Todorov E & Jordan MI ( 2002 ). Optimal feedback control as a theory of motor coordination . Nat Neurosci 5 , 1226 – 1235 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Wilmer JB & Nakayama K ( 2007 ). Two distinct visual motion mechanisms for smooth pursuit: evidence from individual differences . Neuron 54 , 987 – 1000 .
- CrossRef ,
- PubMed ,
- CAS ,
- Web of Science®
- Wyatt HJ & Pola J ( 1981 ). Slow eye movements to eccentric targets . Invest Ophthalmol Vis Sci 21 , 477 – 483 .
- PubMed ,
- CAS ,
- Web of Science®
- Yan YJ , Cui DM & Lynch JC ( 2001 ). Overlap of saccadic and pursuit eye movement systems in the brain stem reticular formation . J Neurophysiol 86 , 3056 – 3060 .
- PubMed ,
- CAS ,
- Web of Science®