MOVIMENTI OCULARI DEL SISTEMA DEL MOVIMENTO OCULARE SACCADICO O RAPIDO
SISTEMA SACCADICO


Le saccadi(fig.1) sono movimenti rapidi degli occhi. Oltre alle saccadi “riflesse”, guidate da eventi periferici (per esempio, quando accade qualcosa di nuovo in visione periferica) e alle saccadi “volontarie”, guidate da strategie interne (per esempio, durante l’esplorazione visiva), sono movimenti saccadici le fasi rapide del nistagmo, le piccole saccadi durante i movimenti di inseguimento lento, i movimenti oculari durante la fase REM del sonno e le microsaccadi durante i periodi di fissazione. A differenza dei riflessi oculari compensatori e in parte del movimento di inseguimento lento, i quali controllano la velocità, il  sistema saccadico controlla la posizione dell’immagine retinica nello spazio. Presiede all’orientamento rapido della fovea verso un bersaglio(fig.2) che si trovi nel campo visivo e sul quale si rivolge l’interesse dell’osservatore.La capacità di compiere dei movimenti saccadici è già presente alla nascita, (fig3)però risultano movimenti spesso imprecisi che hanno bisogno di successive correzioni; a partire dal secondo mese di vita diventano sempre più precisi. I movimenti saccadici, o saccadi, sono spostamenti coniugati dei bulbi oculari, molto rapidi [A saccade ( / s ɨ k ɑː d / sə- KAHD , francese jerk) è rapido], sono movimenti volontari e normali dell’occhio che hanno la funzione di spostare nella zona retinica di massima sensibilità, cioè nella fovea, i vari punti importanti dell’ambiente esterno che stiamo osservando, anche oggetti molto piccoli si proiettano sulla periferia retinica. Vengono eseguite in media 3-4 saccadi al secondo. In media, durante la veglia, vengono quindi eseguite circa 150.000 in un giorno.
sistema saccadico controlla la posizione dell’immagine retinica nello spazio. Presiede all’orientamento rapido della fovea verso un bersaglio(fig.2) che si trovi nel campo visivo e sul quale si rivolge l’interesse dell’osservatore.La capacità di compiere dei movimenti saccadici è già presente alla nascita, (fig3)però risultano movimenti spesso imprecisi che hanno bisogno di successive correzioni; a partire dal secondo mese di vita diventano sempre più precisi. I movimenti saccadici, o saccadi, sono spostamenti coniugati dei bulbi oculari, molto rapidi [A saccade ( / s ɨ k ɑː d / sə- KAHD , francese jerk) è rapido], sono movimenti volontari e normali dell’occhio che hanno la funzione di spostare nella zona retinica di massima sensibilità, cioè nella fovea, i vari punti importanti dell’ambiente esterno che stiamo osservando, anche oggetti molto piccoli si proiettano sulla periferia retinica. Vengono eseguite in media 3-4 saccadi al secondo. In media, durante la veglia, vengono quindi eseguite circa 150.000 in un giorno.
Questo sistema genera dei MOVIMENTI CONIUGATI degli occhi detti SACCADICI i quali hanno le seguenti caratteristiche:
– Sono ESTREMAMENTE VELOCI – La loro velocità può raggiungere i 600-700 °/sec. e ciò è molto utile in quanto durante la loro esecuzione la visione viene sospesa per evitare il disturbo causato dal movimento dell’immagine sulla retina.
Sono ESTREMAMENTE RAPIDI – Il movimento dura frazioni di secondo, ed in particolare inizia 0.2 secondi (latenza) dopo l’individuazione del bersaglio e viene portato a termine in circa 0.05 secondi (tempo d’esecuzione).
– Sono di NATURA BALISTICA – Una volta iniziato un movimento di questo tipo può essere corretto con molta difficoltà nel corso del suo svolgimento. Infatti, una volta che il processo neuronale che provoca un saccadico è iniziato, il sistema di controllo non è in grado di generarne un altro prima di 0.2 sec. indipendentemente dal comportamento del bersaglio.
Per l’esecuzione di un movimento saccadico il sistema DEVE CONOSCERE i seguenti parametri:
a) la POSIZIONE DELL’OGGETTO NELLO SPAZIO – Questo è possibile grazie al valore retino-motorio degli elementi retinici.
b) la POSIZIONE DELL’OCCHIO NELL’ORBITA nel momento in cui l’oggetto viene visto. Questo è possibile grazie alla presenza di particolari neuroni, detti “tonici”, che informano costantemente il sistema di controllo sulla posizione degli occhi nell’orbita.
Questi parametri vengono analizzati, quindi il sistema di controllo genera un comando caratterizzato da:
a) Una determinata DIREZIONE – codificata dal gruppo di motoneuroni che vengono eccitati.
b) Una determinata AMPIEZZA – codificata dalla durata della loro scarica (controllo impulsi ampiezza).
Sebbene nella vita quotidiana le saccadi “avvengano” perlopiù in maniera inconsapevole, esse sono sotto il controllo volontario: possiamo sempre decidere quando e dove eseguire una saccade. Non possiamo però decidere “come” eseguire una saccade e, una volta iniziata, non possiamo fermarla o, solo in condizioni molto particolari, modificarne la traiettoria. Si tratta, infatti, di movimenti dalle caratteristiche molto stereotipate che hanno una tipica forma a gradino (figura 6.4). L’occhio accelera bruscamente, raggiungendo il massimo della velocità nella prima metà della saccade, per poi rallentare quasi altrettanto rapidamente. La velocità e la durata sono calcolate automaticamente dal meccanismo di generazione delle saccadi in base all’ampiezza specificata. Per saccadi di ampiezza inferiore a 20°, vi è una relazione lineare tra ampiezza, velocità massima e durata della saccade. Le saccadi possono avere un’ampiezza compresa fra meno di 0.1° (microsaccadi) e quasi 90°. Raramente, tuttavia, l’ampiezza delle saccadi è superiore a 20-25°: per ampie escursioni dello sguardo si ricorre di regola ad un movimento combinato occhi-testa (si veda in proposito Guitton, 1988). Nell’uomo, una saccade di 5° di ampiezza ha una velocità massima di circa 300% e una durata di circa 50 ms. Saccadi molto ampie possono arrivare ad avere velocità massime di 900°/s e durare fino a 200 ms.
Quanto sono accurate le saccadi?
Quanto bene riusciamo a “centrare il bersaglio” al primo colpo?
Saccadi.fig4 Posizione dell’occhio durante una saccade orizzontale tipica (POS) e relativo profilo di velocità (VEL) e accelerazione (ACC). Si noti la forma a gradino e la brusca accelerazione all’inizio e alla fine della saccade. b Una sequenza di saccadi nell’esplorazione visiva. 0, componente orizzontale; V, componente verticale
Le saccadi servono ad acquisire il bersaglio sulla fovea 2; una volta foveato, il bersaglio deve poi essere mantenuto in quella posizione. Abbiamo anche detto che entro 0.15° dal centro della fovea l’acuità visiva è superiore al 90%, quindi la capacità di discriminazione spaziale è superiore a 0.01° (trascurando l’ iperacuità). Potremmo quindi aspettarci che le saccadi siano accurate entro 0.1-0.2°.
Una serie di esperimenti eseguiti fin dai primi anni settanta aveva rivelato che le saccadi orizzontali non sono molto accurate (si veda Becker, 1991, figura 6.5, simboli aperti). Per saccadi centrifughe verso bersagli a più di 5° di eccentricità, più il bersaglio è lontano, più la saccade “arriva corta”: l’ampiezza saccadica copre solo circa il 90% della distanza del bersaglio. Per distanze inferiori a 5°, il fenomeno invece si inverte: le saccadi sono leggermente ipermetri-che rispetto al bersaglio. Per ampiezze intorno a 5° le saccadi sono normometriche, con un errore medio quasi nullo. Da questi dati sembrerebbe dunque che le saccadi di ampiezza superiore a 10° non riescano a portare il bersaglio in una zona foveale dotata di acuità visiva superiore al 50-60% di quella massima. Questo non vuoi però dire che quando ci guardiamo intorno dobbiamo accontentarci di basse prestazioni visive: intanto l’eventuale errore che rimane al termine della prima saccade può essere corretto mediante una saccade secondaria, di piccola ampiezza e breve latenza (anche meno di 100 ms dal termine della saccade primaria), di cui siamo quasi inconsapevoli.
2 Le saccadi non sono sempre necessariamente legate all’acquisizione di un bersaglio visivo sulla fo-vea. Immaginate di assistere ad uno spettacolo pirotecnico. A parte i possibili movimenti di inseguimento lento, à facile che i vostri occhi siano puntati non già su qualche dettaglio della scena, ma sulla zona cen-trale, in modo da poter abbracciare con lo sguardo tutto lo spettacolo: le saccadi servono anche a tri)posi-zionare l’intero campo visivo.
|
10 20 30 40 50 Eccentricità del bersaglio (gradi) |
|
fig. 5 Errore di posizione al termine delle saccadi in funzione dell’eccentricità del bersaglio. La curva con i simboli vuoti si riferisce a saccadi dirette verso un puntino |

La strategia più efficiente, in termini di tempo, per foveare il bersaglio, è effettivamente accoppiare una saccade primaria con una correttiva e non cercare di essere il più accurati possibile mediante un’unica saccade (Jaeobs. 1987). In secondo luogo, spostare lo sguardo con un movimento combinato occhi-testa aumenta l’accuratezza (Guitton e Volle, 1987). Va poi detto che, con esperimenti effettuati in condizioni più naturali, con bersagli continuamente visibili (e non sotto forma di puntini luminosi che si accendono improvvisamente in un ambiente buio) e in presenza di altri oggetti intorno, si è visto che l’errore medio si mantiene costante, ed è di circa 0.5° (fig. 5, simboli pieni, Collewijn, Erkelens e Steinman, 1988). Infine, l’accuratezza delle sacca-di non è un valore assoluto: per migliorare l’accuratezza possiamo sacrificare, aumentandola, la latenza saccadica (Viviani e Swensson, 1982).
La latenza saccadica media, intesa come il tempo che intercorre tra la comparsa in un momento imprecisato di un puntino luminoso in posizione periferica ignota a circa 10° di eccentricità e l’inizio della saccade, è circa 200 ms. Numerosi fattori possono modificare la latenza delle saccadi. Sono fattori che la aumentano: uno stimolo a luminosità molto bassa, l’eccentricità del bersaglio (bersagli a 60° determinano una latenza di circa 230 ms), la presenza del bersaglio in mezzo a distrattori, ed infine eventuali trasformazioni spaziali da interporre fra uno stimolo luminoso che segnala il momento di partenza e il bersaglio finale (per esempio, le cosiddette anti-saccadi, che sono saccadi in direzione opposta a quella dello stimolo). Altri fattori riducono invece la latenza, come la conoscenza del momento di inizio del movimento o della posizione del bersaglio.
I centri sottocorticali che partecipano alla generazione delle saccadi si trovano nel tronco dell’encefalo e nel collicolo superiore. I centri del tronco dell’encefalo sono principalmente la sostanza reticolare paramediana pontina e la sostanza reticolare mesencefalica. Questi due centri “costruiscono” il tipico pacchetto motorio saccadico da inviare ai motoneuroni oculari sulla base delle specifiche della posizione finale che giungono da altri centri nervosi. Nei centri del collicolo superiore avvengono invece alcune importanti trasformazioni visuo-motorie. È stato proposto che, in particolari condizioni di stimolazione caratterizzate da una latenza saccadica molto breve (circa 100 ms, saccadi espresse; Fischer e Weber, 1993), venga attivato un circuito saccadico rapido, una specie di riflesso optomotorio che, oltre a questi centri “di basso livello”, coinvolge la corteccia occipitale. Tale circuito rapido verrebbe attivato quando le risorse attenzionali non sono impiegate nell’elaborazione dello stimolo visivo all’atto di programmare la nuova saccade. Per le altre saccadi, con latenza di 200¬300 ms, sono implicate anche altre aree della corteccia parietale, frontale e temporale che consentono un’integrazione visuo-motoria più elaborata (figura 6.). Lesioni selettive a queste strutture neurali compromettono in diversa misura la capacità di generare movimenti rapidi degli occhi (Fuchs, Kaneko e Scudder, 1985).

fig.6.Per riuscire a far muovere l’occhio bruscamente, i motoneuroni oculari inviano ai muscoli oculari un breve e potente comando motorio (un treno di spike ad alta frequenza, il cosiddetto “impulso d’innervazione”) per vincere la pur piccola inerzia dell’occhio, in modo da imprimergli una grande accelerazione. Una volta raggiunta la nuova posizione, l’occhio deve poi rimanervi. L’impulso è allora rimpiazzato da un segnale tonico (il cosiddetto “gradino”) proporzionale alla posizione dell’occhio nell’orbita (figura 6.7; Fuchs, Kaneko e Scudder, 1985).
Durante un movimento saccadico si verifica il cosiddetto fenomeno della soppressione saccadica, cioè l’inibizione della visione (Volkmann, 1986). Per averne prova, mettetevi di nuovo di fronte allo specchio e guardatevi alternativamente prima un occhio poi l’altro. Per quanto vi sforziate non riuscirete a vedere il movimento dei vostri occhi. Non si tratta semplicemente di eventi troppo rapidi per essere osservati: le saccadi degli altri le vediamo benissimo.
Avrebbe poco senso parlare delle saccadi senza soffermarsi su un altro elemento fondamentale dell’esplorazione visiva. Stiamo parlando delle fissazioni, cioè i momenti fra una saccade e l’altra durante i quali l’occhio è fermo sul bersaglio. Dal punto di vista del controllo motorio, non si tratta di periodi “passivi”, in cui l’occhio, dopo una saccade, è lasciato a sé stesso: un sistema di stabilizzazione dello sguardo entra in funzione al termine della saccade per mantenere il bersaglio sulla fovea (si veda Kowler, 1991). La stabilizzazione sembra essere garantita dal movimento di inseguimento lento, anche se si tratta di un caso particolare, con lo stimolo a velocità zero.
|
|
|||||||||||
|
|||||||||||
|
Fig.7 Registrazione dell’attività elettrica di un motoneurone oculare che innerva il muscolo retto laterale (tracciato in alto) e contemporanea registrazione della posizione dell’occhio (in basso). Le saccadi in direzione temporale (deflessioni verso il basso) sono accompagnate da un breve treno di impulsi ad alta frequenza (l’impulso d’innervazione). La frequenza degli impulsi nervosi durante le fissazioni è invece correlata alla posizione dell’occhio nell’orbita (più l’occhio è in posizione temporale, più elevata è la frequenza). Si può notare che durante le saccadi in direzione nasale questo motoneurone è inibito e, conseguentemente, il retto laterale viene rilasciato: contemporaneamente però il motoneurone del muscolo antagonista (che innerva il muscolo retto mediale) genera l’impulso d’innervazione che ruota l’occhio nell’altra direzione (organizzazione “a briglie per cavalli”) (fonte: Fuchs, Kaneko e Scudder, 1985) |
|||||||||||

Su questo punto, tuttavia, non vi è ancora un accordo definitivo (Luebke e Robinson, 1988), tanto che talvolta si parla delle fissazioni come di un tipo particolare di movimento di stabilizzazione dello sguardo, non riconducibile al movimento di inseguimento lento.
Tuttavia per quanto ci sforziamo di fissare un oggetto, gli occhi non sono però mai perfettamente immobili durante le fissazioni. Movimenti oculari in miniatura si sovrappongono all’azione del sistema di stabilizzazione e si possono riconoscere: 1) un tremore di piccolissima ampiezza; 2) una lenta fluttuazione erratica della posizione dell’occhio (con una velocità media di circa 0.10.2°/s); 3) delle microsaccadi, la cui ampiezza non eccede di solito 0.2°. Nell’insieme, questi micromovimenti determinano una fluttuazione casuale dello sguardo di circa 0.1° intorno al bersaglio (fig .8). Come vedremo più avanti, lungi dall’essere dannosi, i movimenti oculari in miniatura sono molto importanti, perché una piccolissima fluttuazione dell’immagine retinica è addirittura vitale per la visione.
L’importanza delle fissazioni sta nel fatto che costituiscono i periodi “attivi” della scansione visiva, o, con un’analogia un po’ forzata, i momenti in cui l’otturatore di una macchina fotografica è aperto. La loro distribuzione temporale varia in funzione di molti fattori, ma da esperimenti di ispezione di scene naturali si può ritenere che la durata minima sia circa 100 ms, mentre la durata massima dipende dal tipo di attività esplorativa in cui si è coinvolti, sebbene sia raro trovare fissazioni di durata superiore a 1-2 s. La durata tipica delle fissazioni, intesa come moda della distribuzione, è compresa fra 200 e 300 ms (Viviani, 1990).
Durante le fissazioni viene effettuata l’analisi dello stimolo visivo (fase che implica la perdita di rilevanza dell’attuale stimolo foveale e la selezione di un punto di fissazione periferico) e la programmazione della saccade successiva. Sebbene sia stata formulata l’ipotesi che questi sottoprocessi avvengano sequenzialmente, è possibile che vi sia una parziale sovrapposizione temporale fra di loro (Viviani, 1990).
|
|
|
Movimenti oculari in miniatura durante le fissazioni. Cerchio piccolo: distanza fra due coni nera fovea. Il cerchio grande ha un diametro di 0.1r (fonte: Pritchard, 1964) |

I movimenti oculari disgiunti
I movimenti oculari che abbiamo esaminato fino ad ora sono detti coniugati perché il movimento dei due occhi è uguale. Il meccanismo che mantiene la coniugazione è dovuto ad una particolare organizzazione delle connessioni neuronali a livello dei nuclei oculomotori (legge di Hering dell’uguale innervazione). In molti casi i movimenti oculari non sono però coniugati.
Un caso estremo di movimento non coniugato degli occhi è quello del camaleonte, in cui i due occhi sono completamente indipendenti. Nel camaleonte la diplopia, cioè lo sdoppiamento della visione, è un fatto normale; d’altra parte in questo animale non esiste un meccanismo di fusione binoculare che assicuri la vividezza della percezione tridimensionale. E come se il camaleonte avesse barattato un sistema di campionamento del mondo molto flessibile basato su due “telecamere” indipendenti in cambio della perdita della sensibilità tridimensionale basata sulla disparità retinica. In letteratura è riportato almeno un caso di “uomo-camaleonte”: a causa di un forte strabismo con assenza di corrispondenza retinica, sembra che quest’uomo potesse tranquillamente guidare il camion con un occhio e leggere il giornale con l’altro (Walls, 1962)!
Senza dover chiamare in causa organizzazioni visive particolari, movimenti oculari non perfettamente coniugati si trovano anche nell’uomo, anche se naturalmente di entità minore. Se indossate occhiali le cui lenti hanno proprietà ottiche diverse, l’immagine retinica di uno stesso oggetto sarà leggermente diversa nei due occhi. Un movimento oculare perfettamente coniugato avrebbe pertanto conseguenze visive diverse. Per esempio, una saccade potrebbe essere normometrica in un occhio e ipometrica nell’altro occhio. Il sistema nervoso è però in grado di distribuire il comando motorio binoculare in funzione delle specifiche esigenze visive di ogni occhio. Il risultato è un movimento oculare non ben coniugato (Zee e Levi, 1989).
Ma nemmeno in condizioni normali e con bersagli che non richiedono cambiamento di vergenza le saccadi sono perfettamente coniugate: per ampiezze superiori a 5° le saccadi introducono un disallineamento transiente degli occhi di circa 2-3° (la saccade in abduzione è sempre più ampia di quella in adduzione); dopo la saccade rimane poi un piccolo disallineamento residuo di circa mezzo grado (Collewijn, Erkelens e Steinman, 1988). Nonostante ciò, non vi è pericolo di diplopia perché i valori di disparità retinica che consentono la fusione binoculare possono fluttuare di qualche grado (questa è la ragione dell’esistenza della cosiddetta area di Panum): un non perfetto allineamento dei due occhi è quindi ben tollerato.
Infine, un caso di movimenti oculari per definizione non coniugati è quello dei movimenti di vergenza.
Caratteristiche In genere la loro ampiezza massima è di circa 20°. Oltre tale angolazione di solito entra in gioco la rotazione della testa. La loro massima velocità angolare può raggiungere i 900°/s. La latenza di una saccade è, in media, di circa 225 ms. Questo vuol dire che tra la comparsa del bersaglio e l’inizio della saccade passano circa 225 ms.
La main sequence è una relazione che lega tra loro durata e ampiezza della saccade. In generale la velocità della saccade tende comunque a variare per consentire una durata relativamente costante (questo è vero soprattutto per saccadi di ampiezza relativamente ridotta).
Le saccadi hanno traiettorie rettilinee per i movimenti orizzontali, mentre hanno traiettorie curve per quelli verticali e obliqui.
Funzione : Gli esseri umani e molti animali non guardano una scena che rimane fissa e stabile; invece, gli occhi si muovono, individuando le parti interessanti della scena e costruire una mappa mentale, tridimensionale corrispondente alla scena (in contrapposizione alla mappa grafica degli uccelli, che si basa spesso su rilevazione di movimento angolare sulla retina ). Durante la scansione immediate vicinanze o la lettura, gli occhi umani fanno scatti movimenti saccadici e fermare più volte, muovendosi molto velocemente tra ogni fermata. La velocità di movimento durante ogni saccade non può essere controllato; gli occhi si muovono velocemente in quanto sono in grado. [Neil et al 2010] Una ragione del movimento saccadico dell’occhio umano è che la parte centrale della retina la fovea che fornisce la parte ad alta risoluzione di visione è molto piccola gli esseri umani, solo circa 1 ~ 2 gradi di visione, ma svolge un ruolo fondamentale nella risoluzione di oggetti. Spostando l’occhio in modo che piccole parti di una scena possono essere percepiti con maggiore risoluzione , le risorse del corpo possono essere utilizzate in modo più efficiente.
FISIOLOGIA
Controllo motorio
Gli occhi sono tenuti nell’orbita da 3 paia di muscoli, detti extraoculari:
1. Retto mediale e laterale
2. Retto superiore e inferiore
3. Obliquo superiore e inferiore
· I movimenti laterali coinvolgono principalmente il retto mediale e laterale, e parzialmente gli obliqui.
· I movimenti verticali coinvolgono principalmente il retto superiore ed inferiore.
· I movimenti di adduzione e abduzione coinvolgono i retti mediali e laterali.
· Gli altri movimenti composti chiamano in causa in varia misura diversi muscoli a seconda della loro componente principale.
Il sistema di generazione
Nel cervello possono essere individuate 3 aree principali la cui attività è associata all’esecuzione delle saccadi.
· Campi oculari frontali (FEF)
· Collicolo superiore (SC)
· Nuclei oculomotori del tronco encefalico (BSG)
· A livello dei nuclei del tronco encefalico sono presenti due principali classi di neuroni fondamentali per il movimento saccadico:
1. Cellule pause o omnipause
2. Cellule burst Le cellule “pause” sono attive quando gli occhi sono fermi, quindi la fissazione (sistema WHEN).
· Le cellule “burst” sono attive prima e durante la saccade (sistema WHERE)
In entrambi i casi il fatto che la modifica dell’attività neurale preceda la saccade, suggerisce che questa attività ha un ruolo causale e non è solo correlata all’esecuzione saccadica.
Il Collicolo superiore
 Il collicolo superiore (SC) è una delle aree più importanti per la generazione delle saccadi. SC è la struttura filogeneticamente antica, collocata a livello del tetto del mesencefalo, e che svolge un ruolo cruciale nella esecuzione delle saccadi, soprattutto per quelle di tipo riflesso. (risposta involontaria ad uno stimolo, mediata da elementi nervosi, che termina con una risposta). L’output del SC è il segnale di ingresso principale al sistema di controllo per la generazione delle accadi a livello del tronco dell’encefalo.
Il collicolo superiore (SC) è una delle aree più importanti per la generazione delle saccadi. SC è la struttura filogeneticamente antica, collocata a livello del tetto del mesencefalo, e che svolge un ruolo cruciale nella esecuzione delle saccadi, soprattutto per quelle di tipo riflesso. (risposta involontaria ad uno stimolo, mediata da elementi nervosi, che termina con una risposta). L’output del SC è il segnale di ingresso principale al sistema di controllo per la generazione delle accadi a livello del tronco dell’encefalo.
• Invia informazioni ai centri di generazione delle saccadi nel mesencefalo e nel tronco encefalico.
• Riceve informazioni sia dalla retina che dalla corteccia visiva primaria.
• Trasforma le afferenze visive in comandi oculomotori.
• I neuroni del collicolo superiore hanno campi recettivi che corrispondono a regioni del campo visivo, sensibili sia alla presenza di stimoli visivi, che all’esecuzione di movimenti oculari.
Programmazione delle saccadi
La programmazione saccadica è un processo di tipo gerarchico:
1. Nel primo stadio viene deciso “quando” e in che “direzione” si devono muovere gli occhi.
2. Successivamente viene computata l’ampiezza del movimento.
· Tale parametro si basa sulle caratteristiche del target. Il programma può essere modificato se cambiano le caratteristiche del target, ma solo entro una certa finestra critica dall’inizio della saccade (Becker & Jurgens, 1979). Superata tale finestra critica, la saccade sarà eseguita anche se vengono cambiate le caratteristiche del target. Le saccadi, una volta iniziati non possono essere interrotti o modificati. La finestra critica affinché uno stimolo possa modificare la programmazione di una saccade è di circa 70ms prima dell’onset della saccade.
CENTRO PONTINO DELLO SGUARDO E MOVIMENTI SACCADICI
È un’area della formazione reticolare pontina che controlla i movimenti saccadici, in particolare quelli orizzontali.
A questo livello sono stati identificati quattro tipi di neuroni che sono attivi durante i movimenti saccadici:
a) CELLULE “BURST” – Svolgono un ruolo fondamentale nel processo d’inizio del movimento saccadico e nella regolazione della sua ampiezza.
– L’attività pulse consiste in una serie di impulsi in alta frequenza in cellule dette burst cells, che determinano un’energica contrazione dei muscoli agonisti del movimento da effettuare.
– In questo modo si vince l’inerzia dell’occhio e si impone una forte accelerazione al movimento.
– L’attività delle cellule burst “codifica” sia la direzione che l’ampiezza del movimento oculare.
– Sono dei neuroni che si attivano all’improvviso, circa 10 millisecondi prima dell’inizio del movimento.
– Scaricano a raffica durante l’esecuzione del saccade ed il numero d’impulsi è proporzionale all’ampiezza del movimento saccadico (possono arrivare anche a 1000 impulsi al secondo).
– Cessano la loro attività appena prima che l’occhio raggiunga la nuova posizione, e rimangono silenti fino al saccade successivo.
– Possono venire eccitate da afferenze vestibolari, visive e volontarie.

b) CELLULE TONICHE – Hanno il compito di trasmettere segnali sulla posizione degli occhi e sono attive durante l’esecuzione di tutti i movimenti oculari.
– Un cambio di posizione degli occhi viene associata ad una concomitante variazione della frequenza di scarica di tutti i neuroni tonici: alcuni aumentano la loro attività mentre altri la diminuiscono in base alla nuova posizione assunta. In questo modo il sistema nervoso centrale riceve le informazioni sulla posizione degli occhi nelle orbite.
– Durante la fissazione scaricano a frequenza costante; questa frequenza cresce linearmente all’aumentare della rotazione laterale dell’occhio.
– Durante i movimenti saccadici la loro attività passa dal livello costante che aveva prima del movimento, ad un nuovo livello post saccadico con frequenza di scarica più elevata.
c) CELLULE “BURST” TONICHE – Scaricano a frequenza costante durante la fissazione, mentre presentano un’attività a raffica durante i saccadi laterali. Aumentano la loro frequenza di scarica con la rotazione laterale degli occhi.
d) CELLULE “PAUSE” – Questi neuroni normalmente scaricano ad una frequenza abbastanza costante, ma cessano completamente la loro attività durante i movimenti oculari.
– Si ritiene che la loro attività inibisca le cellule “burst”: la cessazione della loro scarica fa si che le cellule “burst” possano determinare l’insorgenza dei movimenti saccadici.
– L’attività step è determinata da cellule cosiddette pause, o omnipause cells. Queste hanno
un’attività tonica che dipende in ogni momento dalla posizione dell’occhio nell’orbita.
– Pochi ms (5-15) prima di un movimento oculare l’attività di questi neuroni si annulla per un breve periodo.
– L’attività ritorna ritmica e costante al termine del movimento oculare,e dipenderà dalla nuova posizione oculare.
– INTERCONNESSIONI TRA I DIVERSI TIPI DI CELLULE
La figura mostra le ipotetiche connessioni sinaptiche stabilite tra i vari tipi di neuroni.
a) Si ritiene che le cellule burst e i neuroni tonici forniscano afferenze dirette ai motoneuroni.
b) Ad occhio fermo i neuroni pause impediscono ai burst di scaricare.
c) Quando un oggetto appare nel campo visivo ed il sistema decide di guardarlo, si ha un’inibizione dei neuroni pause ed una conseguente scarica ad alta frequenza dei neuroni burst.
d) L’occhio comincia a muoversi e quando arriva sul bersaglio le cellule burst terminano di scaricare.
e) La connessione inibitoria tra i neuroni burst e quelli pause viene disattivata, si ha una riattivazione dei neuroni pause ed una nuova inibizione dei burst.
|
|
|


SCHEMA SEMPLIFICATO
Circuiti sinaptici del centro pontino dello sguardo implicati nell’esecuzione di un movimento saccadico verso destra.
a) I neuroni eccitatori “burst” danno inizio al movimento eccitando sia i neuroni del nucleo dell’abducente (per il retto laterale OD) che i neuroni inibitori della formazione reticolare bulbare.
b) Nel nucleo del VI° vi sono degli interneuroni, i cui assoni attraversano la linea mediana e vanno ad eccitare i motoneuroni dell’oculomotore (per il retto mediale 0S).
c) Anche gli assoni degli interneuroni inibitori della formazione reticolare bulbare attraversano la linea mediana; provocano l’inibizione del nucleo dell’abducente di sinistra (per il retto laterale sinistro) e dell’oculomotore di destra (per il retto mediale destro)
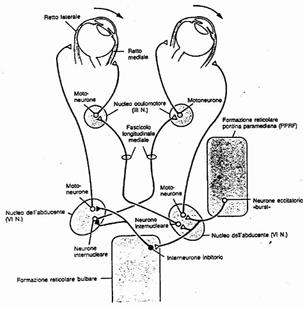 Fig.10
Fig.10
MOVIMENTI OCULARI COINVOLTI NELLA LETTURA
Durante la lettura, gli occhi si spostano continuamente da una parola a quella successiva, per acquisire e quindi elaborare le informazioni contenute nel testo. I movimenti saccadici hanno un ruolo decisivo nella lettura, in quanto permettono agli occhi di spostarsi nei diversi punti della riga ed inoltre di andare a capo alla riga successiva.
Anche la fissazione è importante in quanto, tra un movimento saccadico e l’altro, ci sono delle pause di fissazione in cui avviene il PROCESSO INFORMATIVO, fase in cui avviene la DECODIFICAZIONE DELLE LETTERE che costituiscono le parole.
Per quanto riguarda i movimenti d’inseguimento, questi non intervengono durante la lettura, ma si ritiene che l’abilità nel controllo e nella precisione dei pursuit, sia strettamente correlata alla capacità di eseguire i movimenti saccadici fini richiesti nella lettura
MOVIMENTI SACCADICI
Impiegano approssimativamente il 10% del tempo di lettura.
Hanno un’ampiezza media di 8-9 caratteri che, alla distanza di lettura, corrisponde a 2°.
La loro durata è proporzionale all’ampiezza: per un movimento di 2° sono necessari 25-30 millisecondi, mentre per un movimento di 5° ne servono 30-40.
Durante la loro esecuzione il cervello non è in grado di elaborare nessuna informazione. Questo fenomeno, che prende il nome di SOPPRESSIONE SACCADICA, viene attuato probabilmente per evitare di percepire il movimento e lo sfuocamento dell’ambiente circostante, conseguente al movimento delle immagini del mondo esterno sulla retina, che si verifica durante i saccadi.
Il meccanismo della soppressione saccadica sembra svolgersi a livello della corteccia striata e del collicolo superiore, ed è dovuto principalmente ad una riduzione della sensibilità luminosa.
La soppressione saccadica inizia 40 millisecondi prima del movimento, aumenta progressivamente nella prima parte fino ad annullare la percezione visiva, per poi terminare verso la fine del saccade (vedi figura).

Fig.11
I movimenti saccadici hanno inizio con una stimolazione improvvisa dei muscoli agonisti interessati nello spostamento necessario; contemporaneamente, avviene una completa inibizione degli antagonisti relativi, la quale cessa quando la fovea raggiunge l’oggetto d’interesse.
Nonostante la riattivazione degli antagonisti, può capitare che la fovea oltrepassi l’oggetto, provocando un cosiddetto “overshoot”.
Anche durante la scansione di una riga, possono essere necessari dei movimenti saccadici a ritroso; questi movimenti prendono il nome di “regressioni” e sono necessari per correggere gli overshoot, oppure per rileggere delle parole interpretate male, ma anche quando il soggetto ha delle difficoltà a comprendere il testo. In un lettore normale, le regressioni impegnano dal 5 al 20% di tutta l’attività saccadica svolta durante la lettura.
Antisaccadi (Fig.12)

• Permettono di studiare i meccanismi di pianificazione e controllo delle saccadi.
• Richiedono un alto livello di controllo sul proprio comportamento, e la prestazione spesso contiene diversi “errori”, chiamati prosaccadi.
• Spesso i soggetti non sono consapevoli di aver compiuto queste saccadi nella direzione del target.
• La latenza delle antisaccadi non differisce molto da quella tipica delle saccadi (anche se non si osservano mai “antisaccadi espresse”). Ciò suggerisce che i meccanismi alla base di saccadi e antisaccadi siano diversi e possano operare in parallelo.
Funzioni
Relazioni con l’attenzione
Saccadi e attenzione potrebbero avere una relazione di tipo associativo ma non causale. In altri termini, potrebbe accadere che attenzione e movimenti oculari tendano ad essere diretti nella stessa posizione, ma che i due sistemi non siano necessariamente legati tra loro.
I dati in letteratura sono a favore di una relazione causale tra attenzione e saccadi. Non sarebbe quindi possibile dirigere l’attenzione da un lato e le saccadi in un punto diverso. Comunque, ci sono state parecchie discussioni al riguardo: Klein (1980) hanno fornito evidenze a favore di una possibile dissociazione tra attenzione e movimenti oculari. Una delle evidenze più convincenti del collegamento tra attenzione e saccadi è fornito invece dal lavoro di Hoffman & Subramaniam (1995).
Controllo dei saccadici (fig.13)
 Una volta saccades sono in corso, non possono essere modificati a piacimento. Il cervello non utilizza feedback continuo, dove c’è inseguimento continuo di errore, ma piuttosto risponde alla deriva dell’occhio dal bersaglio intermittenza per ritornare lo sguardo verso il bersaglio; Questo è chiamato un sistema di dati campionati . [Sensory Reception 1987] Un saccade è anche una conseguenza involontaria di rotazione della testa da un lato o l’altro in risposta ad un rumore improvviso da un lato, o un movimento improvviso rilevato nella periferia visiva ( nessun riferimento). Il distacco provocato di saccadi è fatto da tre aree corticali distinte: i campi frontali occhio , i campi occhio parietale , ei campi oculari supplementari . I campi visivi frontali sbloccano la fissazione, e comincia saccadi deliberati di obiettivi visibili o la posizione prevista un oggetto riapparirà in, consentendo l’esplorazione intenzionale della scena visiva. I campi occhio parietale iniziano saccadici riflessivi sulla improvvisa comparsa di bersagli visibili, consentendo l’esplorazione riflessiva della scena visiva. I campi visivi supplementari assistere nell’avvio e nel controllo saccadi fatte durante il movimento del motore (della testa e del corpo), permettendo saccadi per la programmazione a motore complessi. [Pierrot-Deseilligny et al.1995]
Una volta saccades sono in corso, non possono essere modificati a piacimento. Il cervello non utilizza feedback continuo, dove c’è inseguimento continuo di errore, ma piuttosto risponde alla deriva dell’occhio dal bersaglio intermittenza per ritornare lo sguardo verso il bersaglio; Questo è chiamato un sistema di dati campionati . [Sensory Reception 1987] Un saccade è anche una conseguenza involontaria di rotazione della testa da un lato o l’altro in risposta ad un rumore improvviso da un lato, o un movimento improvviso rilevato nella periferia visiva ( nessun riferimento). Il distacco provocato di saccadi è fatto da tre aree corticali distinte: i campi frontali occhio , i campi occhio parietale , ei campi oculari supplementari . I campi visivi frontali sbloccano la fissazione, e comincia saccadi deliberati di obiettivi visibili o la posizione prevista un oggetto riapparirà in, consentendo l’esplorazione intenzionale della scena visiva. I campi occhio parietale iniziano saccadici riflessivi sulla improvvisa comparsa di bersagli visibili, consentendo l’esplorazione riflessiva della scena visiva. I campi visivi supplementari assistere nell’avvio e nel controllo saccadi fatte durante il movimento del motore (della testa e del corpo), permettendo saccadi per la programmazione a motore complessi. [Pierrot-Deseilligny et al.1995]
Tempi e cinematica (fig.14)
 I saccadici so Sono uno dei movimenti più veloci prodotti dal corpo umano ( la rapida chiusura della palpebra può raggiungere velocità di punta ancora più alte). La velocità media (in gradi/sec.) è proporzionale all’ampiezza del movimento. Più ampio è il movimento, > maggiore la velocità MEDIA(es. un movimento saccadico di 60° di ampiezza avrà una vel. di 700° al secondo) La velocità angolare di picco dell’occhio durante una saccade arriva fino a 900 ° / s nell’uomo; in alcune scimmie, la velocità di picco può raggiungere i 1000 ° / s. La latenza dei saccadici ad uno stimolo inatteso normalmente circa 160-200 millisecondi (ms) per avviare e quindi a durata del movimento completo è di 80 – 120 msec., a seconda della loro ampiezza (20-30 ms è tipico in lettura lingua). In determinate circostanze di laboratorio, la latenza, o tempi di reazione per la produzione saccade possono essere tagliati quasi a metà (espresso saccadi).Questi movimenti saccadici sono generati da un meccanismo neuronale che bypassa i circuiti che richiedono molto tempo e attiva i muscoli degli occhi in modo più diretto. [Fischer, B .; Boch, R. 1983 Fischer, B .; Ramsperger, E. (1984)].;). ( ritmi alfa ) specifica pre-bersaglio oscillatorio e le attività transitorie che si verificano nella corteccia parietale posteriore-laterale e nella corteccia occipitale che è anche caratterizzare delle saccadi espresse. [Hamm, JP et al.,2010];
I saccadici so Sono uno dei movimenti più veloci prodotti dal corpo umano ( la rapida chiusura della palpebra può raggiungere velocità di punta ancora più alte). La velocità media (in gradi/sec.) è proporzionale all’ampiezza del movimento. Più ampio è il movimento, > maggiore la velocità MEDIA(es. un movimento saccadico di 60° di ampiezza avrà una vel. di 700° al secondo) La velocità angolare di picco dell’occhio durante una saccade arriva fino a 900 ° / s nell’uomo; in alcune scimmie, la velocità di picco può raggiungere i 1000 ° / s. La latenza dei saccadici ad uno stimolo inatteso normalmente circa 160-200 millisecondi (ms) per avviare e quindi a durata del movimento completo è di 80 – 120 msec., a seconda della loro ampiezza (20-30 ms è tipico in lettura lingua). In determinate circostanze di laboratorio, la latenza, o tempi di reazione per la produzione saccade possono essere tagliati quasi a metà (espresso saccadi).Questi movimenti saccadici sono generati da un meccanismo neuronale che bypassa i circuiti che richiedono molto tempo e attiva i muscoli degli occhi in modo più diretto. [Fischer, B .; Boch, R. 1983 Fischer, B .; Ramsperger, E. (1984)].;). ( ritmi alfa ) specifica pre-bersaglio oscillatorio e le attività transitorie che si verificano nella corteccia parietale posteriore-laterale e nella corteccia occipitale che è anche caratterizzare delle saccadi espresse. [Hamm, JP et al.,2010];
Main sequence saccadici, mostrando singole saccadi da un partecipante l’esecuzione di un compito saccade visivamente guidato.
L’ampiezza di un saccade è la distanza angolare effettuata dall’occhio durante il movimento. Per ampiezze fino a 15 o 20 °, la velocità di una saccade dipende linearmente dalla ampiezza (la cosiddetta sequenza principale saccadica, [Bahill,et al.,1975] un termine mutuato dalastrofisica ; vedi Figura). Per ampiezze maggiori di 20 °, la velocità di picco inizia a plateau [Bahill,et al.,1975] (non lineare) verso la velocità massima raggiungibile dall’occhio a circa 60 °.Per esempio, ampiezza 10 ° è associato a una velocità di 300 ° / s, e 30 ° è associato a 500 ° / s. [Reception sensoriale 1987] Pertanto, per intervalli di ampiezza maggiore, la sequenza principale può essere meglio modellata da un inverso potere legge funzione. [Baloh,et al.,1212]
Le alte velocità di punta e il rapporto sequenza principale può essere utilizzato anche per distinguere tre categorie di Microsaccadi [sono una sorta di movimento oculari di fissazione] ,da altri movimenti oculari come:
a) il tremore oculare (che è una costante, fisiologica, ad alta frequenza (picco 80Hz), bassa ampiezza (stimato circa 150-2500nm (1) b)deriva oculare (movimenti di fissazione degli occhi) e c)movimenti oculari di inseguimento regolari (smooth pursuit ). Basata su Algoritmi di Velocità sono un approccio comune per il rilevamento delle saccade nel movimento degli occhi (eye tracking) . Engbert et al.,2003;[ [11][12][13] Anche se, a seconda delle esigenze di temporizzazione di precisione, i metodi basati sull’accelerazione sono più precisi. [Behrens,et al., 2010 ] [14]
I movimenti Saccadici possono ruotare gli occhi in qualsiasi direzione per spostare la direzione dello sguardo (direzione della vista che corrisponde alla fovea), ma normalmente i saccadici non determinano una rotazione torsionale degli occhi. (Torsione in senso orario o antiorario attorno alla linea di vista quando l’occhio è nella sua posizione primaria centrale; definito in questo modo, la legge di Listing dice che, quando la testa è immobile, la torsione viene mantenuta a zero.)
I saccadici con la Testa-fissa possono avere una ampiezza massima di 90 ° (da un bordo della gamma oculomotor all’altro), ma in condizioni normali i saccadici sono molto più piccoli e ogni spostamento dello sguardo superiori a circa 20 ° è accompagnato da un movimento della testa . Durante tali movimenti saccadici dello sguardo, prima l’occhio produce una saccade per spostare sguardo sul bersaglio, mentre la testa segue tale movimento più lentamente e il riflesso vestibolo-oculare (VOR) Questo riflesso funzioni per stabilizzare immagini sulle retine (in visione aggiogati) durante il movimento della testa producendo movimenti oculari nella direzione opposta al movimento della testa . Dal momento che il VOR può effettivamente ruotare gli occhi attorno alla linea di vista, i movimenti degli occhi e della testa combinati non sempre obbediscono alla legge di Listing. [ citazione necessaria ]
Tipi
|
Peripherally (visually guided) saccades Pro-saccades Express saccades |
Involontarie, riflesse, visive |
|
Centrally guided saccades Predictive saccades (anticipatorie) Memory guided saccades (remembered saccades) Anti-saccades |
Volontarie, finalizzate, modificabili con l’allenamento e la volontà |
Peripherally guided Saccades (pro-saccades) :
Saccadi involontarie, automatiche, riflesse, con tempi di latenza ridotti
Il paziente esegue il movimento in modo involontario o saltuariamente volontario per la comparsa di un oggetto di interesse
 Sono saccadi “guidati dalla vista”(visual guided saccades))
Sono saccadi “guidati dalla vista”(visual guided saccades))
Nessun meccanismo predittivo

Express Saccades:
saccadi talmente rapide da manifestarsi con tempi di latenza vicini ai minimi ritardi di conduzione dell’impulso nervoso afferente ed efferente…..
si suppone siano innescate dalla diretta trasformazione del segnale visivo entrante in comando motorio che guida gli occhi immediatamente verso lo stimolo !!!!
riflesso di “afferramento visivo”
![]()
|
Time reaction after a new retinal visual target |
Saccade time |
(secondary saccade for head-eyes interaction |
Total |
|
160-200 msec. |
80-90 msec |
(200 msec) |
260 to 500 msec.!!!!!!!! |
 From 40 msec before the saccadic movement, the visual suppression is almost complete and the vision becomes normal again only after the stop of the eye movement (in total also for 100 msec).
From 40 msec before the saccadic movement, the visual suppression is almost complete and the vision becomes normal again only after the stop of the eye movement (in total also for 100 msec).
Blindness 4 hour x day!!!!!!!!
The cognitive processes allow the reconstruction of the fluency our life.
vestibular deficit, the patiens are blind for a longer time
change blindness
(cecità al cambiamento)
La change blindness accade durante la soppressione saccadica ma anche se vi sono :
• un campo visivo oscurato o segnale transiente globale(“flicker”)
• altri elementi aggiuntivi o segnali transienti locali (“mud-splashes”)
• l’ammiccamento ad occhi chiusi (“attentional blink”).
Per primi, dopo 200 ms, si muovono gli occhi ( saccade in direzione del bersaglio)
A causa dell’inerzia, la testa si muove 20-50 ms più tardi.
Al termine della saccade oculare la testa sta ancora muovendosi. Durante questa fase gli occhi si spostano in direzione opposta al movimento della testa per compensarne, almeno parzialmente il movimento e l’inerzia
L’angolo fra fovea e posizione extra-foveale inizialmente occupata dall’oggetto è definito errore retinico, La velocità delle saccadi è molto elevata, ne]l’ordine di 700 gradi di arco/sec. Benché l’inizio della saccade sia volontario, la sua velocità dipende dall’errore retinico. Una volta iniziata, la saccade si svolge in modo involontario, e durante questo movimento la visione è soppressa. Raggiunto il bersaglio visivo, il movimento si arresta. Latenza. velocità ed accuratezza sono i parametri che vengono valutati, Questi movimenti, originati dai campi oculari dei lobi frontali, vengono regolati dalla formazione reticolare mesencefalica (saccadi verticali) e pontina (saccadi orizzontali), e controllati dal cervelletto prima di raggiungere i nuclei oculomotori. Disordini cerebellari del verme e dei nuclei fastigiali producono anormalità saccadiche denominate dismetrie, caratterizzate da una scarsa accuratezza dell’arresto del movimento sul bersaglio (“over-shoot” o ipermetria. “under-shoot” o ipometria) (Fig. 1). Dismetrie disconiugate consistenti in ipermetria in un occhio ed un movimento scivolato (glissadico) nell’altro si osservano tipicamente nell’oftalmoplegia internucleare. mentre rallentamenti delle saccadi sono frequenti negli anziani per l’effetto di farmaci del snc. o per malattie muscolari, Alterazioni più marcate. fino all’aprassia possono essere associate a disordini corticali o del troncoencefalo.
La fase rapida del nistagmo può essere considerata un movimento saccadico involontario. che seguendo immediatamente la fase lenta ha la funzione di riportare l’occhio al centro dell’orbita.
Tecniche di acquisizione
Tecniche di acquisizione
Il movimento coniugato saccadico viene evocato da un bersaglio luminoso puntiforme, posto ad un metro di distanza dal paziente e che si accende alternativamente a destra ed a sinistra rispetto alla posizione di sguardo diretto.
La traccia E.OG (di aspetto simil-rettangolare) è caratterizzata dall’ampiezza del movimento, dalla frequenza di alternanza e dal tempo di persistenza del bersaglio nelle singole posizioni. Il bersaglio può muoversi secondo una sequenza regolare con parametri fissi (Figura 7A) o secondo una sequenza random a parametri variabili casualmente nel tempo (Figura 7B).
 FIGURA 7 Registrazione EOG di movimento saccadico normale: bersaglio in movimento con sequenza regolare e parametri fissi (A);
FIGURA 7 Registrazione EOG di movimento saccadico normale: bersaglio in movimento con sequenza regolare e parametri fissi (A);
bersaglio in movimento casuale nel tempo con sequenza random e parametri variabili (B)
Parametri misurati
I parametri più importanti per la valutazione clinica del tracciato EOG sono:
— la velocità angolare del movimento rapido;
— l’accuratezza della parte finale del movimento nel raggiungere il bersaglio.
Nei soggetti normali il movimento saceadico è correttamente eseguito fino a velocit5 angolari del bulbo oculare intorno ai 400°/sec. La velocità angolare di spostamento è in rapporto soprattutto con l’ampiezza del movimento. Nella pratica clinica usiamo per lo più sequenze regolari con frequenza di alternanza di 0,5 Hz ed ampiezze di ± 10° e ± 15°.
METODI DI MISURAZIONE DEI MOVIMENTI OCUIARI3
L’elettrooculografia (EOG)
All’inizio del secolo, con i ricercatori impegnati a registrare potenziali elettrici da ogni possibile e immaginabile parte del corpo, divenne chiaro che degli elettrodi posti in vicinanza degli occhi registravano segnali legati ai movimenti oculari. Tali segnali derivano da una differenza di potenziale di circa 1 mV tra il fundus dell’occhio e la cornea (potenziale corneo-retinico). Tale potenziale si può registrare con un elettrodo posto sull’esterno dell’occhio (per i movimenti orizzontali) e uno sopra o sotto le palpebre (per i movimenti verticali). È la tecnica usata comunemente in clinica. Pur essendo semplice ed economica, tale tecnica però ha numerosi svantaggi. Il principale è che non conosciamo la natura del potenziale corneo-retinico, che non è costante e risente di numerosi fattori, alcuni dei quali non ben chiariti.
Mediante ,alcuni accorgimenti con 1’EOG si possono eseguire registrazioni dei movimenti oculari in un ambito di quasi 30° (sul piano orizzontale, molto meno su quello verticale), con una accettabile linearità e una risoluzione di circa 1°. È comunque sconsigliabile per misurazioni precise.
![]()
![]()
![]() I metodi fotoelettrici
I metodi fotoelettrici
Questi metodi utilizzano degli emettitori di luce infrarossa puntati Ssull’occhio con dei corrispondenti rivelatori della luce diffusa (non riflessa) dall’occhio. Gli emettitori e i rivelatori, con la relativa circuiteria, sono generalmente inseriti in montature simili a occhiali o in una mascherina intorno agli occhi. Il metodo fotoelettrico si basa sul contrasto di luce al limbo (il bordo fra la sclera chiara e l’iride più scuro). È pertanto più facile registrare i movimenti orizzontali che quelli verticali perché i limbi superiore ed inferiore sono facilmente coperti dalle palpebre. Nella forma più semplice basta un unico emettitore di un fascio circolare di luce centrato sul limbo e un rivelatore posizionato in modo da catturarne la luce diffusa, la quale sarà proporzionale alla posizione dell’occhio, a cui corrisponde una diversa proporzione di area illuminata sull’iride e di area illuminata sulla sclera_ Montaggi con più emettitori e rivelatori aumentano la precisione di misura e sono quindi da preferirsi. È importante che nella registrazione fotoelettrica si minimizzino o si compensino le fluttuazioni di luce ambientale, dato che le componenti infrarosse che giungono sull’occhio modificano il segnale. Una fonte di disturbo è costituita dall’alternanza a 50Hz della luce nelle lampadine ad incandescenza. È molto meglio usare un’illuminazione a fluorescenza. Un altro problema consiste negli spostamenti della montatura rispetto alla testa. Dato il raggio oculare (in media 13 mm), uno spostamento di soli 0.1 mm comporta un artefatto nel segnale corrispondente ad una rotazione di circa 0.5°. È quindi fondamentale, anche se di non facile attuazione (anche solo per l’elasticità della pelle), rendere perfettamente stabile la montatura. Con un buon montaggio si possono ottenere registrazioni lineari in un ambito di circa 30° per i movimenti verticali e di circa 40° per quelli orizzontali. La precisione può arrivare a quasi 0.1°. Il limite massimo di frequenza è piuttosto elevato e senza troppe difficoltà si può arrivare a 1000Hz.
La videooculografia
Un fascio di luce proiettato sull’occhio produce un’immagine riflessa, detta immagine primaria di Purkinje (la classica finestra distorta negli occhi di alcuni personaggi dei fumetti). Questa è dovuta alla superficie esterna riflettente della cornea. Poiché il centro di rotazione dell’occhio non coincide con il centro di curvatura della cornea, quando l’occhio si muove, l’immagine riflessa cambia posizione. Tale cambiamento di posizione può essere registrato mediante una telecamera che riprende l’occhio. Si possono anche utilizzare alcune caratteristiche della pupilla (quali area, centroide, forma) per migliorare il sistema. Per minimizzare i segnali dovuti alla traslazione dell’occhio si possono usare le immagini secondarie di Purkinje, dovute alle riflessioni della superficie interna della cornea e delle superfici interna ed esterna del cristallino. Il grande vantaggio di questa metodica è che non è richiesta alcuna montatura. È quindi più “ecologica” rispetto ad altre tecniche. Tuttavia è costosa e complicata da mettere a punto, e con un limite massimo in frequenza imposto dalla tecnologia video (attualmente vi sono telecamere a 200 o addirittura 500 Hz, che sono valori elevati, ma spesso il limite è 50 Hz).
II metodo della bobina sclerale
Tale metodo si basa sul principio per cui in una bobina immersa in un campo magnetico si genera un potenziale elettrico che è funzione del piano della bobina rispetto alla direzione del campo magnetico. Il campo magnetico è generato da una coppia di bobine di grandi dimensioni. Il soggetto sta al centro del campo magnetico e indossa una speciale lente a contatto al cui interno è immersa la bobina in miniatura che genera il segnale. La bobina è quindi solidale con i movimenti dell’occhio.
Questo è il metodo più accurato per registrare i movimenti oculari, sebbene richieda che i soggetti indossino la lente a contatto. La precisione nominale può giungere a 0.01° e virtualmente non vi sono limiti nella frequenza massima. Si candida come il miglior metodo in laboratorio, ma è difficilmente utilizzabile per altri scopi, quali rassessment clinico, le indagini di marketing o l’uso quale interfaccia uomo-macchina (portatori di handicap, consolles virtuali e così via).
L’osservatore umano
Annoverare l’osservatore umano fra gli strumenti di misurazione dei movimenti oculari è volutamente provocatorio. La provocazione è duplice. Da un lato, risulta infatti che siamo strumenti migliori, in termini di precisione della “misura”, di alcune tecniche come l’EOG: l’uomo è in grado di discriminare la direzione dello sguardo di un’altra persona, in condizioni ottimali, con una precisione superiore al grado (Cline, 1967)! D’altra parte, quello di paragonarci a degli strumenti di misura è evidentemente un pretesto che ci consente di richiamare l’attenzione su una capacità molto importante, ossia la nostra capacità di capire dove sta guardando un’altra persona e la possibilità di stabilire o meno il contatto visivo (“guardarsi negli occhi”). È del tutto superfluo qui ricordare in quali e quanti contesti di comunicazione interpersonale la direzione dello sguardo è un fattore fondamentale del comportamento sociale, financo nel regno animale
MOVIMENTI OCULARI
Nell’esame clinico del paziente la parte più importante riguarda l’esame dei movimenti oculari, ed in particolare la ricerca del nistagmo. Il nistagrno è definito come una serie di movimenti oculari (scosse) ciascuno costituito da una fase lenta che sposta il centro del bulbo verso la periferia, seguita da una fase rapida che riporta il bulbo al centro. La fase rapida denomina la direzione del nistagmo. Questo fenomeno se osservabile con capo fermo ed in posizione normale indica quasi sempre un disordine del sistema vestibolare e più precisamente un’alterazione del normale riflesso vestibolo-oculo motore (VOR).
Il VOR benché non richieda uno stimolo visivo, è tuttavia costantemente interattivo con gli stimoli visivi, e ciò ne aumenta notevolmente l’efficienza. Il VOR è necessario per stabilizzare la visione foveale durante i movimenti della testa. evitando così l’offuscamento delle immagini. I meccanismi centrali dell’oculomozione1)sistema di inseguimento oculare lento, 2)sistema saccadico. sistema otticocinetico. ed in misura minore il sistema di vergenza, combinano i loro effetti con il VOR per ottenere istante per istante la più nitida rappresentazione visiva del mondo circostante. Pertanto il VOR viene classificato entro la famiglia dei movimenti oculari che costantemente permettono di ottenere una visione distinta.
Nei paragrafi successivi si riporta una sintetica descrizione dei sistemi che presiedono ai vari tipi di movimento oculare classificati nella Tabella I,
CLASSIFICAZIONE MOVIMENTI OCULARI
|
Definizione |
Stimolo |
Funzione |
|
Inseguimento lento |
Bersagli lenti |
Fovea su bersagli visivi mobili |
|
Saccadici |
Bersagli rapidi |
|
|
Vergenza |
Bersagli frontali a distanza variabile |
|
|
VOR |
Piccoli movimenti del capo |
Fovea su bersagli mobili con capo in movimento |
|
Otticocinetici |
Successioni di bersagli o rotazione sostenuta ad occhi aperti |
|
Tab I. Cinque sistemi neuronali partecipano a mantenere la visione foveale Sistema di inseguimento lento mantiene l’immagine sula fovea mentre il bersaglio è in movimento. Sistema saccadico dirige rapidamente la fovea sul bersaglio che appare nella periferia del campo visivo. Sistema di vergenza muove i bulbi oculari in direzione opposta, cosicché la stessa immagine si proietta su entrambe le fovee (bersagli in movimento avanti-indietro). Sistema vestibolo-oculomotore dirige i bulbi oculari in direzione opposta a rotazioni brevi e veloci (2-4 Hz) della testa.Sistema ottico-cinetico’ usa informazioni visive per stabilizzare la fovea durante rotazioni della testa lente e prolungate
Test oculomotori: [ Il sistema dei movimenti oculari di inseguimento lento (smooth pursuit);il sistema saccadico ; movimenti di vergenza oculare; Il riflesso vestibolo oculo-motore; Il sistema otticocinetico ]
I movimenti d’inseguimento lento e saccadici possono essere facilmente studiati sotto videonistagmografia e/o videooculografia.. Essi permettono in pratica otorinolaringoiatrica di esplorare in pochi minuti in maniera non invasiva le vie oculogire e quindi le vie vestibolari centrali. Ultimamente si è assistito ad una rivalutazione dei movimenti oculari pursuit e saccadici nella pratica otoneurologica
anni 80 : lo studio dei movimenti oculari facevano parte integrante dell’esame vestibolare di II° livello(ENG);per il loro contributo topo diagnostico e per il monitoraggio di malattie neurologiche e terapie associate Per provocare movimenti oculari coniugati di inseguimento, al fine di studiare lo smooth pursuit, e saccadici possono essere utilizzate diverse tecniche di stimolazione che hanno in comune un input visivo caratterizzato da un bersaglio luminoso puntiforme in movimento lento su di un piano che coincide con quello dell’asse visivo diretto alternativamente verso un lato e l’altro con un movimento pendolare per il pursuit . La registrazione vera e propria dei saccadici si effettua facendo eseguire movimenti di fissazione di un led che si accende alternativamente con frequenza ed ampiezza random) sul piano orizzontale Il paziente, seduto ad una distanza di i metro dallo schermo, fissa il punto luminoso, in genere una “barra di LED”. La prova viene condotta in ambiente oscuro e silente. Il movimento oculare e lo spostamento del punto luminoso vengono separatamente registrati con ENG,

Fig. 15 – Movimento di inseguimento lento (tracciato superiore: movimento oculare; tracciato inferiore: movimento della mira).
A) movimenti di inseguimento normali; B) nello spostamento degli occhi verso destra, il movimento di inseguimento è sostituito da piccoli movimenti saccadici di rifissazione.

Fig. 16 – Vari tipi di anomalie dei movimenti saccadici (tracciato superiore: movimento oculare; tracciato inferiore: movimento della mira).
A) saccadi normali; B) dismetria saccadica (OD e OS tracciati superiori): i saccadici sono sostituiti da movimenti irregolari che raggiungono il target la cui fissazione non riesce ad essere mantenuta per tendenza dell’occhio a scivolare in posizione primaria. Microsaccadi di rifissazione per mantenere la fissazione del target o per correggere saccadi ipometriche; C) dismetria saccadica (poliradicoloneurite): ridotto spostamento dei bulbi oculari con tendenza allo scivolamento verso la posizione primaria di sguardo.
anni 90 Viene utilizzato lo stesso sistema di mire luminose o un proiettore LCD, soltanto che il sistema di registrazione ENG /VNG è dotato di apparecchiatura computerizzata per l’elaborazione delle tracce ottenute
|
|
|
|
|
|






anni 00 Fig. 17a-b-c-d-e
Il test impulsivo cefalico (head impulse test – HIT)fig. 18 a-b-c- fornisce una valutazione rapida e precisa, specifica per l’orecchio, del riflesso oculo-vestibolare (VOR) in risposta a stimoli nell’intervallo delle alte frequenze, l’intervallo naturale degli spostamenti del capo dandoci informazioni sui 3 canali semicircolari. fig. 19a-b


Un paziente con perdita vestibolare bilaterale
|
|
|


Test Impulsivo Cefalico ICS – Video Impulse test (vHIT)

fig. 18 a-b-c- paziente con ESAME DEI TRE
CANALI SEMICIRCOLARI

Grafico dei 3 Canali Semicircolari d/s Snapsys) Video testa Impulse Test (VHIT Fig. 19a-b
Il sistema oculare saccadico provvede con movimenti assai rapidi (700°/sec.) a correggere la posizione degli occhi in modo che l’immagine di un oggetto, presente nel campo visivo, venga a cadere sulla fovea. Oltre che da uno stimolo visivo o acustico, il movimento oculare saccadico può venir determinato da uno stimolo sensitivo sia tatti- le che dolorifico. Nell’uomo, la maggior parte dei movimenti saccadici hanno una ampiezza media di circa 15°. La saccade è un movimento oculare del tipo più frequente. Consiste in rapidi movimenti degli occhi eseguiti per portare la zona di interesse a coincidere con la fovea. Vengono eseguite in media 3-4 saccadi al secondo. In media, durante la veglia, vengono quindi eseguite circa 150.000 in un giorno. Sono un movimento di tipo coniugato.
● E’ un movimento volontario, che si osserva durante l’esplorazione della scena visiva.
● Permette di portare nuovi oggetti potenzialmente interessanti sulla fovea.
● E’ un movimento di tipo balistico: una volta iniziato non può esserne cambiata la velocità né la traiettoria.
● La latenza media è di 150-200 ms, dura circa 50 ms e può raggiungere la velocità di 900°/sec.
● L’informazione più importante per l’esecuzione di una saccade è la posizione verso la quale deve essere compiuto il movimento.
● La maggior parte delle volte le saccadi non raggiungono subito il bersaglio, ma devono essere
corrette da una saccade secondaria. ● Se è necessario eseguire saccadi più precise, si noterà un aumento della latenza, che riflette il tempo necessario per programmarle.
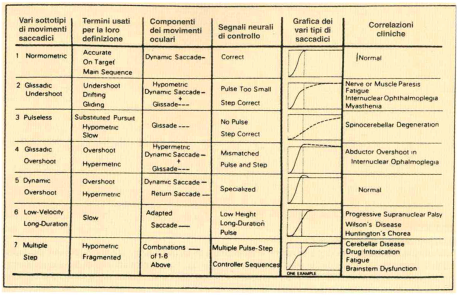
Colonna 1: sottotipi di movimenti saccadici. Colonna 2: termini comunemente usati nella definizione dei saccadici. Colonna 3: componenti dei movimenti oculari. Colonna 4: segnali neurali di controllo. Colonna 5: simulazione al computer dei movimenti oculari che mostra la posizione degli occhi in funzione del tempo. Colonna 6: correlazioni cliniche. La linea continua rappresenta la componente del saccadico dinamico; la linea tratteggiata rappresenta la componente glissadica; la linea punteggiata rappresenta altre componenti dei movimenti oculari come, per esempio, la fase di ritorno di un sorpasso rapido dinamico (da Bahill TA. e ColI., 1979).
Caratteristiche
In genere la loro ampiezza massima è di circa 20°. Oltre tale angolazione di solito entra in gioco la rotazione della testa. La loro massima velocità angolare può raggiungere i 900°/s. La latenza di una saccade è, in media, di circa 225 ms. Questo vuol dire che tra la comparsa del bersaglio e l’inizio della saccade passano circa 225 ms.
La main sequence è una relazione che lega tra loro durata , ampiezza, velocità di picco (Bahill, 1975) della saccade. A questa si aggiungono la latenza (tempo intercorso fra la comparsa del target e l’inizio dello spostamento degli occhi) e l’accuratezza (rapporto posizione degli occhi rispetto al target: guadagno saccadico). La velocità degli occhi aumenta rapidamente fino a circa la metà del saccadico poi diminuisce. Vi è una correlazione stretta fra la velocità massima raggiunta e l’ampiezza del movimento stesso. La relazione fra ampiezza della saccade e durata è lineare. In generale la velocità della saccade tende comunque a variare per consentire una durata relativamente costante (questo è vero soprattutto per saccadi di ampiezza relativamente ridotta).
Le saccadi hanno traiettorie rettilinee per i movimenti orizzontali, mentre hanno traiettorie curve per quelli verticali e obliqui.
Latenza saccadica
La latenza (normale intorno ai 200. msec.) è influenzata da numerosi fattori legati allo stimolo: (luminanza, contrasto, distanza dal punto di fissazione precedente) o dall’età e dalla collaborazione del paziente, ma può essere espressione di specifiche patologie (aprassie oculomotorie).
• E’ il tempo che intercorre fra la comparsa di un target (su cui il soggetto deve portare lo sguardo) e l’inizio della saccade.
• Rappresenta il tempo richiesto dai meccanismi cerebrali che mediano l’orientamento visivo.
• Può essere influenzata da diverse caratteristiche relative sia allo stimolo bersaglio che al contesto sperimentale in cui si trova il soggetto.
• La latenza saccadica varia in funzione della posizione dello stimolo: maggiore è l’eccentricità e maggiore tende ad essere la latenza.
• E’ influenzata inoltre dall’intensità dello stimolo. Stimoli più facilmente discriminabili (ad es. con maggiore contrasto rispetto allo sfondo) richiedono latenze saccadiche minori
L’accuratezza può avere diverse alterazioni talora anche fisiologiche come l’affaticamento.
Lo squilibrio, fra i segnali centrali che determinano il movimento saccadico da luogo a dismetrie: glissade, saccadici ipometrici o ipermetrici (Tab. 3)
Premesse di Anatomo-Fisiologia
Come già sopra riportato i movimenti saccadici servono per portare il più rapidamente possibile lo sguardo su un bersaglio in modo che la sua immagine cada sulla fovea. Se il bersaglio è visivo, la sua identificazione è compito del sistema visivo, mentre la programmazione del movimento oculare è affidata alla corteccia cerebrale. Le aree coinvolte nel controllo corticale dei saccadici sono quelle del lobo parietale, per i saccadici riflessi (per esempio verso un bersaglio che compare improvvisamente nel campo visivo), e quelle del lobo frontale, per i saccadici volontari (per esempio verso un bersaglio già presente nel campo visivo, o verso la posizione memorizzata di un bersaglio, o verso un bersaglio che si prevede che comparirà nel campo visivo), e la Dorso-Lateral Prefrontal Cortex (DLPFC), che inibisce l’esecuzione involontaria di movimenti riflessi (per esempio verso un bersaglio che compare improvvisamente nel campo visivo ma che non riveste alcuna importanza). Le aree corticali proiettano alle strutture premotorie del troncoencefalo, in parte direttamente e in parte indirettamente attraverso il collicolo superiore; quest’ultimo, in rapporto soprattutto con i saccadici volontari, è inibito tonicamente dalla pars reticulata della substantia nigra, che a sua volta è sotto il controllo del nucleo caudato, attraverso una via diretta, che la inibisce, liberando il collicolo superiore, e una via indiretta che la eccita, passando attraverso il nucleo subtalamico di Luwis.
I campi oculari frontali (FEF, frontal eye fields) ( Fig. 20).
|
|
• Ricevono input dalle aree parietali e frontali. • Inviano output al collicolo superiore. • Hanno cellule che rispondono alla presenza di stimoli visivi nel campo recettivo. • Contribuiscono a formare la mappa di salienza del campo visivo. • La risposta di una cellula agli stimoli visivi presenti nel proprio campo recettivo aumenta se questi sono il bersaglio di una saccade. • Diminuisce se gli stimoli sono distrattori che devono essere ignorati. • Possono mostrare selettività per attributi (colore, forma) che contraddistinguono il target, in un dato contesto sperimentale. |

Il controllo sottocorticale dei saccadici è affidato essenzialmente ad un gruppo di neuroni premotori (burst neurons e neuroni omnipausa), al cervelletto e al collicolo superiore. I burst neurons (BN) nel complesso costituiscono il burst generator, il cui compito è quello di generare il segnale di velocità (pulse), che consente di vincere le forze visco-elastiche dell’orbita per spostare gli occhi verso il bersaglio; i neuroni omnipausa (OPN) hanno invece il compito di inibire tonicamente i neuroni del burst generator, che essendo molto instabili potrebbero scaricare spontaneamente anche quando non è necessario, Il cervelletto attraverso il verme dorsale esercita un’azione inibitoria sulle cellule omnipausa, controllando la durata dell’attivazione dei neuroni del burst generator e quindi l’accuratezza del saccadico; per mezzo del lobo flocculo-nodulare, invece, controlla l’integrazione del segnale di pulse, in modo che possa essere generato un segnale di step di durata e di ampiezza adeguate. Il collicolo superiore infine rappresenta un’importante stazione sia per l’avvio dei saccadici che per il controllo della loro metrica. Nel troncoencefalo si riconoscono due popolazioni di BN, gli short leading burst neurons (SLBN) e i long leading burst neuron (LLBN).
Gli SLBN veicolano il segnale di velocità per i movimenti saccadici, e possono essere di tipo eccitatorio (ESLBN) o inibitorio (ISLBN), connessi rispettivamente con i motoneuroni oculari agonisti e antagonisti. Nel caso dei saccadici orizzontali, gli ESLBN sono localizzati nella Formazione Reticolare Paramediana Pontina (PPRF) e sono direttamente eccitatori per il muscolo abducente ipsilaterale; gli ISLBN si trovano invece nella Formazione Reticolare midollare (MedRF) e sono direttamente inibitori per il muscolo abducente controlaterale. Per i saccadici verticali e torsionali, gli ESLBN si trovano nel nucleo rostrale interstiziale del Fascicolo Longitudinale Mediale (riMLF) e sono indirettamente eccitatori, tramite interneuroni, per i muscoli oculomotore e trocleare ipsilaterali, mentre gli ISLBN sono localizzati nel nucleo interstiziale del Cajal (INC) e sono indirettamente inibitori, anche in questo caso tramite interneuroni, per i muscoli oculomotore e trocleare controlaterali. Gli SLBN hanno la caratteristica di scaricare circa 10 msec prima del movimento oculare; la loro frequenza di scarica, la durata e il numero di impulsi sono funzione rispettivamente della velocità, della durata e dell’ampiezza dei saccadici,
I LLBN veicolano il commando per la metrica dei saccadici, e fanno da tramite tra il collicolo superiore e gli SLBN. I LLBN sono localizzati nella porzione più rostrale della PPRF, nella Formazione Reticolare mesencefalica centrale (cMRF) e in altre parti del tronco come il ponte, nel nucleo reticolare del tegmento del ponte (NRTP), ma anche nel midollo rostiale e nel collicolo superiore. Tali neuroni posseggono un’attività di scarica che compare alcune centinaia di msec prima del burst legato al saccadico Nei LLBN avvengono due trasformazioni del segnale proveniente dal collicolo superiore: la riorganizzazione dell’output del collicolo superiore in termini di direzione delle linee di forza dei muscoli oculari e la conversione della codificazione spaziale del collicolo superiore nella codificazione temporale degli SLBN, ( Fig. 21/A).
|
|
Fig. 21/A (a) Sezione parasaggittale del tronco cerebrale di scimmia rhesus. L’area ombreggiata nella parte più alta del tronco cerebrale corrisponde alla Formazione Reticolare Mesencefalica (MRF) nella quale è compreso il NrMLF. La PPRF è localizzata a livello della parte rostrale del ponte. Sia il NriMLF che la PPRF contengono burst cells che codificano l’informazione sulla direzione ed ampiezza dei movimenti saccadici. La parte mediano-caudale della PPRF contiene le pause cells che giocano un ruolo di coordinazione di tutti i movimenti saccadici. (b) Rappresentazione schematica delle vie neuronali sc = collicolo superiore; PC = commissura posteriore; TR = tractus retroflexus; nD = nucleo di Darkschewitsch; ppH = nucleo preposito dell’ipoglosso; Med RF = formazione reticolare midollare; MLF = fascicolo longitudinale mediale; iC = nucleo interstiziale di Cajal; III = nervo oculomotore; NIII =nucleo dell’oculomotore; IV = nervo trocleare; NIV =nucleo del trocleare; VI = nervo abducente; NVI = nucleo dell’abducente; NVII = nucleo del faciale. (modificata da U. Büttner, T. Brandt – Oculor motor disorders of the brain stem – in “Clinical Neurology”, Vol. 1, Nr. 2, August 1992, Baillière Tindall, London).
|

Anche gli OPN si trovano nel troncoencefalo, in prossimità della linea mediana del ponte, nel nucleo del rafe interposto, rostralmente al nucleo dell’abducente, Tali cellule inviano efferenze agli SLBN, sui quali esercitano, come abbiamo visto, un’azione tonica inibitoria, e ricevono afferenze dai neuroni trigger del collicolo superiore, che a sua volta blocca le cellule omnipausa quando deve cominciare il saccadico. Infatti, gli OPN sono collegati agli SLBN eccitatori e inibitori sia pontini che mesencefalici, pertanto è possibile ipotizzare un’inibizione tonica sulle cellule burst per far cessare un saccadico. In maniera inversa quando è richiesto un saccadico gli OPN devono essere inibiti per permettere la scarica dei BN. Quindi gli OPN scaricano continuamente eccetto che immediatamente prima e durante il saccadico e, se vengono stimolati durante un saccadico, il movimento cessa. Poiché la loro attività è presente per movimenti saccadici di qualsiasi direzione sono stati definiti come omnidirectional pause neurons, da cui il termine di neuroni omnipausa. ( Fig. 21/B).
|
|
Fig. 21/B Relazione esistente fra Pause cells (P), Burst cells (B), ed Integratore Neurale (NI) nella generazione del pulse e dello step nel movimento saccadico. Le Pause cells cessano di scaricare prima di ogni saccade, consentendo alle Burst cells di generare il pulse. Il segnale di pulse viene integrato dall’integratore neurale per la produzione dello step. La combinazione del pulse e dello step determina una variazione di scarica sui motoneuroni oculari (OMN) che genera il movimento saccadico dell’occhio (E). Le linee verticali parallele rappresentano l’attività di ciascun neurone (spikes), mentre i grafici mostrano la relazione esistente fra il tempo e l’entità della scarica neuronale. (modificata da R.J. Leigh, D.S. Zee – The neurology of eye movements -2nd ed., F.A. Davis Co., Philadelphia 1991).
|

L’input che guida i saccadici è fornito dai BN eccitatori (EBN) ipsilaterali, che proiettano sia sui motoneuroni che sugli interneuroni dell’abducente ipsilaterale, e sui BN inibitori (IBN) ipsilaterali: a loro volta, i primi proiettano sul retto laterale ipsilaterale, i secondi proiettano attraverso il Fascicolo Longitudinale Mediale (FFM) controlaterale ai motoneuroni per il retto mediale del nucleo del terzo controlaterale, e gli IBN ipsilaterali inibiscono sia il nucleo del sesto controlaterale che gli FBN e gli IBN controlaterali. In questo complesso circuito gli OPN proiettano si. tutti e quattro i gruppi di BN, inibendoli tonicamente, eccetto che durante il saccadico (Figura 5).
|
|
Fig. 22 – La generazione dei saccadici orizzontali I saccadici richiedono un’innervazione reciproca per i muscoli retti mediale e laterale (RM e RL) di entrambi gli occhi Il RL di destra è comandato dai motoneuroni (MN) del nucleo dell’abducente (VI n) ipsilaterale Il nucleo del VI contiene anche un interneurone (IN) che manda le sue proiezioni attraverso il fascicolo longitudinale mediale (FLM) al nucleo del III controlaterale, sui MN che comandano il RM dell’altro occhio Quindi gli EBN forniscono il comando ai MN e agli IN ipsilaterali. Gli EBN proiettano anche agli BN ipsilaterali Gli IBN inibiscono i MN e gli IN controlaterali Cosi la coppia EBN/IBN fornisce un’innervazione reciproca ai muscoli retti. Gli IBN inibiscono anche gli EBN e gli IBN controlaterali. Una conseguenza di questo cross-coupling è che le coppie EBN/ IBN di entrambi i lati formano un circuito a feedback positivo a breve latenza Quando i neuroni omnipausa (OPN) sono attivi impediscono a questo circuito di essere instabile.
|

Il cervelletto, abbiamo detto, partecipa al controllo dei saccadici, da una parte controllandone la metrica attraverso il verme dorsale, dall’altra contribuendo al meccanismo di integrazione del segnale di pulse in quello di step con il lobo flocculo-nodulare. Un’altra area del cervelletto correlata con i movimenti saccadici è rappresentata dal nucleo del fastigio caudale, i cui neuroni scaricano all’inizio dei saccadici controlaterali per accelerarli e alla fine dei saccadici ipsilaterali, per frenarli (Fig.23).
|
|
Fig.23 – Diagramma semplificato del burst generator troncoencefalico per i saccadici orizzontali (da A.F.Fuchs et al, 1993). Le direzioni contro(laterale) e ipsi(laterale) sono relative al nucleo del fastigio (FN) di sinistra.Lo spessore della linea che parte dal FN indica la forza di quella proiezione sulla base dell’interpretazione che gli autori hanno dato ai dati anatomici presenti in letteratura (Noda et al, 1990). Gli schemi dei patterns di scarica dei neuroni del FN si riferiscono ai saccadici verso destra e verso sinistra. FN, neurone del nucleo del fastigio; IBN, neurone burst inibitorio; EBN, neurone burst eccitatorio; OPN, neurone omnipausa; VI, motoneurone abducente.
|

Rimane da considerare il collicolo superiore, che rappresenta la sede fondamentale per dare inizio al movimento saccadico e per controllarne la metrica una volta che questo è cominciato. Il collicolo superiore è costituito da più strati di neuroni, di cui quelli dello strato superficiale sono sensibili all’input retinico, scaricano 80 msec dopo la comparsa del target visivo, hanno una rappresentazione retinotopica e rispondono solo alla posizione dello stimolo visivo. Ci sono poi altri neuroni, localizzati negli strati profondi, anch’essi correlati con il movimento oculare. Infine, ci sono cellule che presentano una scarica sostenuta quando lo stimolo è nel campo recettoriale della cellula, che continua a scaricare finchè il saccadico non allontana lo stimolo dal campo recettoriale e si attivano in modo che la loro localizzazione nella mappa motoria corrisponda al vettore di errore di posizione del saccadico. Nel collicolo superiore, inoltre,
si riconoscono due zone funzionalmente importanti, una connessa con il controllo della fissazione, posta più cranialmente e che si attiva durante la fissazione, un’altra, che prepara il movimento saccadico, posta più caudalmente e che si attiva in occasione del movimento.
Durante la fissazione di un target, si attivano i neuroni collicolari connessi con la fissazione in entrambi i poli rostrali, mentre i neuroni collicolari restanti sono inibiti; dopo l’apparizione di un target nel campo visivo, si attiva un insieme di neuroni collicolari posti caudalmente e la cui posizione nella mappa motoria definisce l’errore di posizione dello sguardo. La zona caudale aumenta progressivamente la sua attività a spese del collicolo craniale, che viene depresso. Con il decremento dell’attività di fissazione si verifica un ulteriore aumento dell’attività del collicolo caudale e questo porta all’attivazione dei LLBN. Quando poi la zona rostrale di fissazione è completamente inibita, vengono bloccati i neuroni omnipausa, e questo porta all’inizio del movimento saccadico.
Con il progredire del movimento diminuisce la distanza tra target e fovea per cui si modifica la distribuzione spaziale della zona neuronale attiva nel collicolo, che si muove progressivamente verso il polo rostrale; avvicinandosi al polo rostrale diventano attivi i neuroni collicolari connessi con la fissazione, si attivano di nuovo i neuroni omnipausa, che sbioccano i BN e il movimento saccadico si arresta immediatamente. Da quanto abbiamo detto, risulta chiaro che per generare un saccadico è necessario attivare il burst generator, affinché venga prodotto un segnale di velocità, ma bisogna anche mettere a riposo le cellule omnipausa, in modo che i burst neurons possano essere attivati. Inoltre, il movimento dei due occhi deve essere coniugato e a questo provvedono degli interneuroni che dal nucleo del nervo abducente di un lato proiettano, attraverso il fascicolo longitudinale mediale controlaterale, al nucleo del nervo oculomotore dell’altro lato, al sottonucleo per il muscolo retto mediale. Infine, gli occhi devono essere in grado di rimanere sul bersaglio una volta che lo hanno raggiunto; per questo c’è bisogno anche di un segnale di posizione, per vincere le forze di ritorno elastico dei muscoli oculari, che tenderebbero a riportare gli occhi al centro dell’orbita. Il segnale di step viene generato da un altro gruppo di neuroni, quelli del gaze holding neural integrator, che attraverso un’operazione matematica di integrazione trasforma il segnale di velocità in un segnale di posizione. Tale struttura è localizzata per i saccadici orizzontali nel nucleo preposiro dell’ipoglosso e nel nucleo vestibolare mediale e per i saccadici verticali, oltre che su questi nuclei, anche sul nucleo rostrale interstiziale del FLM e sul nucleo interstiziale del Cajal.
Fisiologia
Controllo motorio
Gli occhi sono tenuti nell’orbita da 3 paia di muscoli, detti extraoculari:
1. Retto mediale e laterale
2. Retto superiore e inferiore
3. Obliquo superiore e inferiore
· I movimenti laterali coinvolgono principalmente il retto mediale e laterale, e parzialmente gli obliqui.
· I movimenti verticali coinvolgono principalmente il retto superiore ed inferiore.
· I movimenti di adduzione e abduzione coinvolgono i retti mediali e laterali.
Gli altri movimenti composti chiamano in causa in varia misura diversi muscoli a seconda della loro componente
Programmare una saccade
Si ipotizza che esistano tre meccanismi che operano in parallelo, coinvolti
rispettivamente nello scegliere:
– Quando (when) sarà eseguita la saccade
– Dove (where) sarà diretta la saccade
– Cosa (what) sarà il bersaglio della saccade
Movimento saccadico Fig.24a/b
|
|
I movimenti saccadici vengono generati da un’attività di tipo pulse-step dei motoneuroni posti in alcune regioni del nel tronco encefalico, che costituiscono la cosiddetta brainstem saccade generator region: – MRF, midbrain reticular formation, – PPRF, paramedian pontine reticular formation.
|


Movimento saccadico
Veloce movimento oculare (“movimento balistico”) che permette di dirigere rapidamente lo sguardo verso una mira.
Comprende:
saccadi semplici (volontarie o involontarie)
fase rapida del Ny vestibolare ed otticocinetico
movimenti oculari rapidi della fase REM
movimento “cieco”
dubbia natura riflessa
“bottom up” e “top down” strategia (saccadici indotti dalla visione
e saccadici guidati centralmente)
Il sistema di controllo delle saccadi comprende:
Lobi parietali
FEF
Collicolo superiore
Cervelletto: verme dorsale (lobuli V, VI e VII), nucleo del
fastigio
Talamo
Gangli basali
Il movimento saccadico è composto da:
una fase “esplosiva” p (burst ‐ saccadic pulse) che genera la forza necessaria per vincere la componente viscosa oculare e per muovere velocemente l’occhio
una fase tonica (saccadic step) che mantiene l’occhio nella nuova posizione contro le forze elastiche di ritorno
Il generatore del “saccadic pulse” si trova nella formazione reticolare pontina paramediana e comprende:
I neuroni Burst di tipo eccitatorio ed inibitorio, a media e lunga conduzione, che scaricano solo durante la saccade
I neurons Pause, che scaricano continuamente eccetto immediatamente prima e durante la saccade (inibiscono tonicamente le burst cells)
I neuroni Tonici, che scaricano in relazione alla posizione oculare alla fine della saccade (mantengono l’occhio nella nuova posizione)
Il sistema di generazione
Nel cervello possono essere individuate 3 aree principali la cui attività è associata all’esecuzione delle saccadi.
· Campi oculari frontali (FEF)
· Collicolo superiore (SC)
· Nuclei oculomotori del tronco encefalico (BSG)
· A livello dei nuclei del tronco encefalico sono presenti due principali classi di neuroni fondamentali per il movimento saccadico:
1. Cellule pause o omnipausa
2. Cellule burst
· Le cellule “pause” sono attive quando gli occhi sono fermi, quindi la fissazione (sistema WHEN).
· Le cellule “burst” sono attive prima e durante la saccade (sistema WHERE)
In entrambi i casi il fatto che la modifica dell’attività neurale preceda la saccade, suggerisce che questa attività ha un ruolo causale e non è solo correlata all’esecuzione saccadica.
Il Collicolo superiore
Il collicolo superiore (SC) è una delle aree più importanti per la generazione delle saccadi. SC è la struttura filogeneticamente antica, collocata a livello del tetto del mesencefalo, e che svolge un ruolo cruciale nella esecuzione delle saccadi, soprattutto per quelle di tipo riflesso. L’output del SC è il segnale di ingresso principale al sistema di controllo per la generazione delle saccadi a livello del tronco dell’encefalo.
Programmazione delle saccadi
La programmazione saccadica è un processo di tipo gerarchico:
1. Nel primo stadio viene deciso “quando” e in che “direzione” si devono muovere gli occhi.
2. Successivamente viene computata l’ampiezza del movimento.
· Tale parametro si basa sulle caratteristiche del target. Il programma può essere modificato se cambiano le caratteristiche del target, ma solo entro una certa finestra critica dall’inizio della saccade (Becker & Jurgens, 1979). Superata tale finestra critica, la saccade sarà eseguita anche se vengono cambiate le caratteristiche del target. Le saccadi, una volta iniziati non possono essere interrotti o modificati. La finestra critica affinché uno stimolo possa modificare la programmazione di una saccade è di circa 70ms prima dell’onset della saccade.
Funzioni
Relazioni con l’attenzione
Saccadi e attenzione potrebbero avere una relazione di tipo associativo ma non causale. In altri termini, potrebbe accadere che attenzione e movimenti oculari tendano ad essere diretti nella stessa posizione, ma che i due sistemi non siano necessariamente legati tra loro.
I dati in letteratura sono a favore di una relazione causale tra attenzione e saccadi. Non sarebbe quindi possibile dirigere l’attenzione da un lato e le saccadi in un punto diverso. Comunque, ci sono state parecchie discussioni al riguardo: Klein (1980) hanno fornito evidenze a favore di una possibile dissociazione tra attenzione e movimenti oculari. Una delle evidenze più convincenti del collegamento tra attenzione e saccadi è fornito invece dal lavoro di Hoffman & Subramaniam (1995).
In questo esame, il compito è quello di seguire un bersaglio che effettua spostamenti bruschi, regolari o aleatori. I parametri misurati sono: la velocità massima o media del movimento saccadico; la latenza tra la partenza del movimento del bersaglio e quella dell’occhio, in teoria vicina a 250 ms; la precisione del movimento saccadico che è il rapporto tra l’ampiezza del movimento dell’occhio e l’ampiezza del movimento del bersaglio. Le dismetrie saccadiche (precisione anomala del movimento saccadico) sono facilmente individuabili su un grafico che indica i valori normali (precisione in funzione della velocità dei movimenti saccadici).
Aspetti cimici e strumentali delle alterazioni dei saccadici
Nello studio EOG dei movimenti saccadici, le alterazioni estrinseche da Ny vestibolare risultano di scarso rilievo clinico: la presenza di Ny (che non si inibisce sotto fissazione) può interessare solo parzialmente il plateau della traccia EOG; alterazioni marcate si riscontrano di solito solo in presenza di Ny congenito. Di notevole importanza sono invece le alterazioni intrinseche al sistema saccadico: nella Tabella IV Sono schematizzati i principali tipi di patterns FOCI patologici per i movimenti saccadici.
TABELLA IV – Le alterazioni dei movimento saccadico
|
A) ALTERAZIONI ESTRINSECHE |
C) METODOLOGIA DEI TESTS SACCADICI |
|
Ny vestibolare – Ny sul plateau |
D) DISTURBI DELLA VELOCITÀ DEI SACCADICI 1. Troppo lento 2. Troppo veloce |
|
E) DISTURBI DELLA LATENZA (TIMING) |
|
|
B) ALTERAZIONI INTRINSECHE |
F) ACCURATEZZA
|
|
– ipermetria (overshoot) |
1Overshoot
|
|
– frenage (undershoot) |
2Undershoot
|
|
– dismetria disconiugata |
3Cecità
|
|
– guadagno ridotto |
4Glissades
|
|
– errori di precisione |
4Pulsion
|
|
– mancata evocazione |
(oscillazioni saccadiche con latenza tra ogni saccade) |
L’ipermetria saccadica (dismetria overshoot) la saccade orizzontale iniziale è troppo grande e la saccade correttivo avviene nella direzione opposta allo spostamento di destinazione, è caratterizzata dal superamento del bersaglio da parte dell’occhio con successi o centraggio per mezzo di un movimento correttivo di ritorno Si parla di sorpasso dinamico glissato (Fig.25A in alto) quando il movimento di correzione è immediato, di tipo lento e di sorpasso dinamico statico (Fig. 25B al centro) quando l’occhio persiste sul bersaglio per breve tempo prima di eseguire il movimento di correzione, L’ipermetria saccadica è espressione di patologia archicerebellare e risulta di solito ipsilaterale (o comunque prevalente) al lato della lesione. E di frequente riscontro nei tumori e negli infarti cerebellari: nella patologia espansiva dell’angolo ponto-cerebellare in stadio neuro-chirurgico, la dismetria oculare è di solito un segno clinico precoce rispetto alla dismetria degli arti.
Il frenage saccadico (under-shoot, Fig. 25C in basso) è rappresentato da un movimento oculare a normale velocità angolare nella fase di avvicinamento al bersaglio, con successivo rallentamento nella fase finale di raggiungimento del bersaglio.

Fig. 25 Traccia EOG di MS con ipermetria saccadica [overshoot]: a sorpasso dinamico glissato A (in alto), a sorpasso dinamico statico B (al centro). Frenage saccadico [undershoot] C (in basso) [da Pagnini – Patterns EOG visuo – oculomotori … in “Potenziali evocati uditivi…”, C.R.S. Amplifon, MI, 26 – 27/11/82].
|
|
|
Fig. 25 D: Dismetria Orizzontale per superamento. In blu è rappresentato la mira luminosa (bersaglio), in verde il movimento orizzontale dell’occhio, in rosso in basso il movimento verticale. L’occhio spesso overshootta il bersaglio. Questo paziente aveva una cavernoma cerebellare centrale ed un peduncolo di spurgo a sinistra che è stato sottoposto poi a resezione. Non vi è alcun superamento per saccadi verticali |

La Figura Fig. 25 D mostra una dismetria overshoot in un paziente con una lesione cerebellare.
Anch’esso è espressione di patologia archicerebellare: manifestazioni di ipometria cerebellare si riscontrano abbastanza spesso nelle eredo-atassie e nelle oftalmoplegie plus.
Talvolta l’ipermetria ed il frenage saccadico Sono presenti nei due occhi in forma disconiugata: l’occhio abdotto esegue un movimento ipermetrico e contemporaneamente si osserva un’ipometria nell’occhio addotto; tale comportamento è presente in entrambe le direzioni del movimento saccadico (Fig. 26). Il reperto è patognomonico di oftalmoplegia internucleare se la convergenza oculare e conservata. L’oftalmoplegia internucleare si manifesta per una lesione dei neuroni internucleari ed è quasi sempre espressione di una sclerosi multipla.

Fig. 26-Registrazione EOG di movimento saccadico in presenza di oftalmoplegia internucleare In occhio destro (rosso) si apprezza l’ipermetria saccadica mentre in occhio sinistro (verde) si evidenzia il frenage (ipometria) [da Pagnini – Patterns EOG visuo – oculomotori … in “Potenziali evocati uditi‑vi…”, C.R.S. Amplifon, MI, 26 – 27/11/82).
Se la velocità angolare di esecuzione del movimento risulta inferiore a 250°-300°/sec (Fig. 27A), si può parlare di riduzione patologica del guadagno neurale del sistema saccadico. Un deficit di guadagno si può riscontrare in varie forme patologiche del tronco encefalico, soprattutto a livello pontino (eredo-atassie, tumori intrinseci, emi sindromi vascolari) ed è quasi sempre presente in forma marcata nelle oftalmoplegie plus. Errori di precisione sul bersaglio si possono riscontrare anche in alcuni soggetti normali (Fig. 27B), : si deve pertanto attribuire un significato patologico solo alla presenza di evidenti e ripetuti movimenti di correzione.

Fig. 27 – Registrazione EOG di movimento saccadico con guadagno ridotto con ipometria (in alto)A ed errori di precisione ipermetria sacca dica (in basso) B [da Pagnini – Patterns EOG visuo – oculomotori … in“Potenziali evocati uditivi…”, C.R.S.Amplifon, MI, 26 – 27/11/82].
La Dismetria Overshoot non è sempre anormale. Nei soggetti normali, transitori overshoot dismetrici sono comuni nelle saccadi rivolte in posizione primaria, nelle saccadi con movimento inferiori a circa 10 gradi , e nelle saccadi fatte ad uno stimolo che appare in una posizione romanzo. I soggetti normali, tuttavia, sapranno riaggiustare le loro saccadi per una posizione di destinazione prevedibile e, dopo diversi refissazioni allo stesso luogo, smetteranno di produrre superamenti. Dismetria Overshoot è anormale quando è frequente (almeno il 50% del tempo), di dimensioni significative (maggiore di 2 gradi), e quando si verifica in saccades centrifughe superiori a 20 gradi.
|
|
|
|
Fig.2 vista assiale e sagittale del cervelletto con tumore metastatico dal seno coinvolge il verme. Il tumore è l’area irregolare bianca al centro di ciascuna immagine. Questo paziente aveva una profonda dismetria saccadica. |
|

|
|
|
Fig.3 Dismetria saccadica in pazienti con lesioni del Verme (immagine cortesia della Dr. Dario Yacovino ). |

![]() Materiale supplementare sul sito DVD : Video della dismetria saccadica (stesso paziente come sopra).
Materiale supplementare sul sito DVD : Video della dismetria saccadica (stesso paziente come sopra).
Nell’aprassia di Cogan e nelle paralisi progressive sopra nucleari il movimento saccadico può risultare anche del tutto abolito. Le paralisi sopra nucleari esordiscono per lo più con una impossibilità ad eseguire movimenti saccadici sul piano verticale; successivamente tali movimenti non sono più possibili nemmeno sul piano orizzontale; di solito in questa patologia i disturbi funzionali dei saccadici precedono quelli dei movimenti di pursuit. Una patologia intrinseca del sistema saccadico si può spesso associare a segni di compromissione del sistema vestibolooculomotorio. In presenza di ipermetria saccadica si riscontra spesso una iperreflessia vestibolare alle prove caloriche e talvolta un Ny di rimbalzo ed un Ny laterale bidirezionale. Nelle ipometrie saccadiche si può ritrovare un Ny calorico che, all’EOG, presenta apici arrotondati. Con una certa frequenza, la dismetria oculare è in associazione con una aritmia del Ny calorico. La dismetria oculare disconiugata delle oftalmoplegie internucleari è sempre accompagnata da un Ny di lateralità dissociato (presente solo nell’occhio abdotto). In altri casi, alterazioni del Ny vestibolare e del Ny ottico-cinetico sono del tutto giustificate da una compromissione primitiva a carico del sistema saccadico. Un ridotto guadagno neurale saccadico comporta necessariamente un Ny ottico-cinetico demoltiplicato ed una iporeflessia vestibolare per deficit di elaborazione centrale del Ny calorico. Se il movimento saccadico non è più evocabile, durante la stimolazione ottico- cinetica e termica si assiste ad uno scivolamento dei bulbi oculari in posizione laterale, con assenza quindi di scosse nistagmiche. La completa compromissione del generatore neurale tronco-encefalico per i movimenti saccadici è quasi sicuramente alla base della mancanza della fase rapida del Ny calorico nell’aprassia di Cogan e nei comi pontini.
C) Metodologia del test di calibrazione:
Il messaggio da portare a casa principale è che il software ITA commerciale è terribile a saccadi, e si deve guardare le tracce, e l’occhio del paziente, e utilizzare il proprio giudizio clinico. Ci sono molti rari disturbi saccades – bisogna avere un sacco di conoscenze per fare molto di loro, e non pagare che spesso non ce ne sono molti di questi pazienti.
Il problema tecnico è che saccades sono molto veloci, e produttori ENG generalmente hanno ottimizzato i loro prodotti per le cose con calma (come nistagmo calorico). Frequenze di campionamento di 15-30 / secondo vanno bene per calorics, ma semplicemente non fanno molto bene per qualcosa che si muove a 600 gradi / secondo. Si consideri – a 600 gradi / sec, l’occhio poteva andare a 30 gradi in 0,05 secondi. Questo è un campione a 20 / sec. E ‘generalmente accettato che bisogna provare almeno a 100 / sec per avere più di una possibilità di saccades misura. Il video non è solo a questo – – perché il frame rate usuale è solo 30 / secondo. Esso non può essere fatto (con telecamere standard).
Inoltre, i produttori ENG non hanno fatto un ottimo lavoro a costruire un software affidabile che quantifica le misurazioni saccadici. Noi abitualmente ignoriamo la grafica di sintesi sul nostro sistema ENG riguardanti la velocità e la latenza, perché fanno tanti errori.Ci basta guardare le tracce (che possono trarre in inganno anche) e il monitor.
I dispositivi che misurano i movimenti oculari sono discussi qui .
|
Tabella 1: Metodologie di registrazione di movimento degli occhi |
Misurano |
Non misurano |
|
EOG (elettrooculografia) |
|
La Torsione oculare |
|
IR (a raggi infrarossi) |
Bene gli orizzontali, male i verticali |
La Torsione oculare |
|
Video |
Orizzontale e verticale moderatamente bene |
La Torsione solo mediante ispezione o usando, attrezzature insolite non disponibili in commercio (dal 2004) |
|
Coil occhio sclerale |
Horizonal e verticale molto bene |
Con misure particolari in grado di misurare la torsione moderatamente bene |
Panoramica:
Test saccadici volte è utile come disturbi cerebellari e disturbi degenerative del sistema nervoso centrale, a volte può essere rivelati attraverso test saccadici. Mentre l’esame clinico da un esaminatore esperto è il metodo più efficiente per rendere questi diagnosi, tali competenze non è sempre disponibile. I tre parametri saccadici più rilevanti per i medici sono picco di velocità , la latenza , e la precisione .
D) Disturbi della Velocità dei saccadici
|
Tabella 2: velocità di picco di 20 saccades deg nei soggetti normali |
|
|
|
METODO |
Velocità di picco |
Limite inferiore di normalità |
|
Infrarossi (300 di larghezza di banda hz) |
657 |
491 |
|
Bobina Eye (60 banda hz) |
650 |
325 |
|
EOG (15 h larghezza di banda) |
336 |
252 |
I valori normali per la velocità di 20 gradi saccades sono riportati nella tabella 2. Si noti che la velocità è molto sensibile al metodo con cui saccades vengono registrate. Normali valori di velocità saccadici ottenuti tramite metodi infrarossi o sclerali registrazioni bobina di ricerca sono in genere superiori a quelli ottenuti con le registrazioni EOG. Metodi EOG e video attualmente predominano registrazioni oculomotori. Questi due metodi hanno larghezze di banda simili e limiti così simili per normale. Queste non sono solo molto bravi a tutti per il test saccadici.
Velocità saccadico è approssimativamente proporzionale all’ampiezza saccadico per dimensioni tra 5 e 20 gradi. Dopo ampiezza raggiunge i 20 gradi, la velocità saccadica subisce una saturazione morbida rispetto a ulteriori aumenti di ampiezza. Questo modello è visto su sequenze principali , che velocità di picco complotto contro formato saccade. Il limite superiore al solito per la velocità saccadici, non importa quanto grande la saccade, è di circa 750 ° / sec. L’autore utilizza la funzione data dall’equazione 1 (vedi sotto) per i suoi limiti di velocità normale. Per il limite inferiore, l’asintoto è impostata a 350 ° / sec. Per il limite superiore, l’asintoto è 750 ° / sec. Ampiezza saccade è indicato con E e la velocità saccadica, E puntino. Velocità saccadici non può essere alterato volontariamente e non è influenzato sostanzialmente dall’età o dal sesso.
Eqn 1
![]()
Ci sono vari problemi di cui essere consapevoli nel misurare velocità saccadici. Variabilità è apprezzabile e uno si consiglia di acquistare circa 40 saccades di dimensioni variabili tra i 10 ei 40 gradi per sviluppare una ragionevole sequenza principale. Errore di calibrazione è un altro problema comune. L’errore di taratura può essere correlato a fattori sottili, che non sono evidenti quando il test oculomotore viene letto. Ad esempio, i pazienti con paralisi motorie oculari sono in grado di ottenere uno o entrambi gli occhi al bersaglio. Pazienti con strabismo possono alternare l’occhio che vedono da, a seconda della direzione dello sguardo, e permettere un occhio alla deriva fuori dal bersaglio. In questi casi, velocità saccadici misurati sono sbagliate, perché la taratura è inadeguato. Registrazione monoculare e la visualizzazione solo occhio sono essenziali per evitare errori in questo tipo di pazienti. Nei pazienti senza problemi di allineamento oculare, la prova che la calibrazione è stabile per diversi studi deve essere disponibile prima diagnosi di anomalie del saccades.
D)a. Saccadi lente:
D)b. Saccades Too Fast
Saccades anormalmente veloci
• Errore di calibrazione
• Opsoclono
• Sindromi di restrizione
Anormalmente saccades veloci di solito possono essere ricondotti a un errore di calibrazione o una registrazione rumoroso movimento degli occhi. Questi possono essere molto difficili da individuare apparecchiature ENG come commerciale è di solito in grado di produrre molte informazioni utili in materia saccades troppo veloci (o eccessivamente quelle lente o).
Errori di calibrazione:
|
|
Errore di calibrazione in un paziente con INO che faceva saccadi aspetto più normale di quanto non siano in realtà. La traccia superiore mostra che l’escursione occhio è molto più grande della destinazione. La traccia in basso mostra che mentre saccades adduzione sono più lente, tuttavia essi sono molto vicini alla normalità. Se si dovesse ridurre le velocità in proporzione all’errore calibrazione, non sarebbe normale nella registrazione di questo paziente. Così, il test di laboratorio è fuorviante. Questa registrazione è stata realizzata con Micromedical |

Come esempio di errore di calibrazione, si consideri il caso in cui un paziente effettua una saccade 5 gradi ad uno spostamento di destinazione 15 gradi. Poi fattore di calibrazione sarà tre volte troppo grande, e saccadi apparirà anormalmente veloci. Registrazioni Noisy, come ad esempio a causa di un elettrodo di EOG male applicata o artefatto batter sono un’altra causa di quello che sembra essere saccadi impropriamente veloci. Poiché la velocità saccadico viene calcolato dalla velocità di picco, rumore velocità aggiunge alla velocità di picco reale, e si traduce in modo errato alte velocità. Ciò è particolarmente un problema con le registrazioni infrarossi, che combinano elevata larghezza di banda con suscettibilità a lampeggiare artefatto.
Un esempio più sottile di un errore di calibrazione è mostrato nella figura precedente. Qui, un paziente che era ovviamente anormale nella clinica, sembrava quasi normale sui test oculomotor. Ciò è dovuto a una calibrazione errata che ha fatto tutte le velocità saccade artifactually più grande. Così, una calibrazione sbagliato può fare qualcuno che è molto anomalo, guarda quasi normale!
Errore di calibrazione in un paziente con INO che faceva saccadi aspetto più normale di quanto non siano in realtà. La traccia superiore mostra che l’escursione occhio è molto più grande della destinazione. La traccia in basso mostra che mentre saccades adduzione sono più lente, tuttavia essi sono molto vicini alla normalità. Se si dovesse ridurre le velocità in proporzione all’errore calibrazione, non sarebbe normale nella registrazione di questo paziente. Così, il test di laboratorio è fuorviante.
Questa registrazione è stata realizzata con Micromedical apparecchiature per la tecnologia.
Come esempio di errore di calibrazione, si consideri il caso in cui un paziente effettua una saccade 5 gradi ad uno spostamento di destinazione 15 gradi. Poi fattore di calibrazione sarà tre volte troppo grande, e saccadi apparirà anormalmente veloci. Registrazioni Noisy, come ad esempio a causa di un elettrodo di EOG male applicata o artefatto batter sono un’altra causa di quello che sembra essere saccadi impropriamente veloci. Poiché la velocità saccadico viene calcolato dalla velocità di picco, rumore velocità aggiunge alla velocità di picco reale, e si traduce in modo errato alte velocità. Ciò è particolarmente un problema con le registrazioni infrarossi, che combinano elevata larghezza di banda con suscettibilità a lampeggiare artefatto.
Un esempio più sottile di un errore di calibrazione è mostrato nella figura precedente. Qui, un paziente che era ovviamente anormale nella clinica, sembrava quasi normale sui test oculomotor. Ciò è dovuto a una calibrazione errata che ha fatto tutte le velocità saccade artifactually più grande. Così, una calibrazione sbagliato può fare qualcuno che è molto anomalo, guarda quasi normale!
Opsoclono
 Il termine opsoclono (opsos = visione , klonos = movimento rapido) rappresenta movimenti involontari incontrollati degli occhi, caotici e multivettoriali cioè ad orientamento orizzontale, verticale e diagonale. Questa anomalia, definita anche saccadomania, scompare durante il sonno. Sia nel flutter oculare che nell’opsoclono le saccadi generate risultano incomplete e costituite soltanto dalla porzione pulse mentre manca la parte step, di conseguenza viene a mancare l’intervallo intersaccadico (200ms),che di norma separa 2 saccadi successive. La fisiopatologia delle oscillazioni senza intervallo intersaccadico non è stata ancora chiarita L’ipotesi di Zee e Robinson (1979) di una instabilità funzionale dei neuroni omnipausa non è stata confermata dagli studi sperimentali condotti da Kaneko (1996) Il disturbo potrebbe interessare un’ampia rete neuronale del ponte e del cervelletto, ma al momento non si sa quali neuroni e quali trasmettitori siano colpiti. Più frequentemente del flutter, l’opsoclono si accompagna a vere atassie e a mioclonie di differente gravità, queste ultime sono spasmi asincroni di gruppi muscolari del tronco, delle estremità e del collo (spesso accentuati o scatenati dall’ammiccamento, dall’intenzione al movimento, dagli stati emotivi). La sindrome flutter-opsoclono-mioclono colpisce nel 50% dei casi i bambini al di sotto dei 5 anni e in circa il 50% dei casi è provocata da un neuroblastoma. I rimanenti casi riconoscono una natura idiopatica, para e post-infettiva, postvaccinale, encefalitica. Il quadro clinico dell’adulto è identico, ma presenta sostanziali
Il termine opsoclono (opsos = visione , klonos = movimento rapido) rappresenta movimenti involontari incontrollati degli occhi, caotici e multivettoriali cioè ad orientamento orizzontale, verticale e diagonale. Questa anomalia, definita anche saccadomania, scompare durante il sonno. Sia nel flutter oculare che nell’opsoclono le saccadi generate risultano incomplete e costituite soltanto dalla porzione pulse mentre manca la parte step, di conseguenza viene a mancare l’intervallo intersaccadico (200ms),che di norma separa 2 saccadi successive. La fisiopatologia delle oscillazioni senza intervallo intersaccadico non è stata ancora chiarita L’ipotesi di Zee e Robinson (1979) di una instabilità funzionale dei neuroni omnipausa non è stata confermata dagli studi sperimentali condotti da Kaneko (1996) Il disturbo potrebbe interessare un’ampia rete neuronale del ponte e del cervelletto, ma al momento non si sa quali neuroni e quali trasmettitori siano colpiti. Più frequentemente del flutter, l’opsoclono si accompagna a vere atassie e a mioclonie di differente gravità, queste ultime sono spasmi asincroni di gruppi muscolari del tronco, delle estremità e del collo (spesso accentuati o scatenati dall’ammiccamento, dall’intenzione al movimento, dagli stati emotivi). La sindrome flutter-opsoclono-mioclono colpisce nel 50% dei casi i bambini al di sotto dei 5 anni e in circa il 50% dei casi è provocata da un neuroblastoma. I rimanenti casi riconoscono una natura idiopatica, para e post-infettiva, postvaccinale, encefalitica. Il quadro clinico dell’adulto è identico, ma presenta sostanziali
differenze riguardanti l’eziologia, la terapia, la prognosi. Fino attorno al 40° anno si incontrano prevalentemente forme encefalitiche perlopiù benigne, oltre questa età si equivalgono encefaliti e sindromi paraneoplastiche da carcinoma: bronchiale a piccole cellule, mammella, utero, ovaie, vescica, tiroide (ordinati sulla base della decrescente frequenza ).Il frequente rilievo di anticorpi nucleari antineurali (ANNA-2 e Anti-Ri), soprattutto in corso di carcinomi del seno e di altri tumori ginecologici, denota la genesi immunologica di almeno una parte delle sindromi
opsoclono-paraneoplastiche Sia i flutter che l’opsoclono sono frequenti nella patologia cerebellare, a seguito di encefaliti virali. I movimenti possono avere una piccola ampiezza, appaiono cioè come minuscole deviazioni rispetto alla posizione primaria. In rari casi, saccades veloci in modo anomalo possono essere reali, e non a causa di un artefatto tecnico. Una causa è la opsoclono sindrome o è relativo, battito oculare . In queste condizioni, i pazienti fanno saccadi indesiderati senza intervallo intersaccadico, che possono essere eccessivamente veloce per le loro dimensioni (Bergenius, 1986). Sindrome Kinsbourne o Síndrome de opsoclono-mioclono
l’Opsoclonus denota caotici movimenti oculari saccadici in avanti ed indietro. È una sindrome drammatica, solitamente indica o un cancro o una encefalite del tronco cerebrale. L’Opsoclono differisce essenzialmente dalla flutter , nell’ opsoclono le saccadi sono orientati in qualsiasi direzione (orizzontale, verticale, torsione), mentre nel flutter è generalmente sempre puramente orizzontale. L’Opsoclono può essere difficili da registrare. Si verifica così rapidamente – con una frequenza fino a 20 Hz – che non può essere catturato da dispositivi a bassa ampiezza di banda come gli EOG clinici o i sistemi VNG . La Videoregistrazione può anche avere difficoltà a catturare questo rapido movimento in quanto il frame rate può essere solo 30 fps. L’Opsoclono si può generalmente visualizzare meglio utilizzando un sistema di occhiali video Frenzel , con un grande schermo.
![]() Video di opsoclono in giovane donna, sviluppato dopo una pandemia virale del Nilo occidentale, osservata a Chicago. Vedere il sito DVD pagina per un elenco di altri film come questo.
Video di opsoclono in giovane donna, sviluppato dopo una pandemia virale del Nilo occidentale, osservata a Chicago. Vedere il sito DVD pagina per un elenco di altri film come questo.
L’Opsoclono è quasi sempre causato da un tumore neuronale (a neuroblastoma) o sindrome paraneoplastica (un tumore in altre parti del corpo). Queste diagnosi sono molto rare e di conseguenza opsoclono è raramente visto e ancor più raramente riconosciuto. Un iter in piena regola per neoplasia è indicato e produttivo in persone con Opsoclonus. Il cancro ai polmoni è il tumore comunemente trovato e quindi con una radiografia del torace (piuttosto che un MRI) è solitamente il primo test più produttivo. Recentemente, la PET è stato usato per diagnosticare tumori occulti in questa situazione (Bataller et al, 2003). Anche se gli anticorpi come anti-Hu, Yo e Ri tra gli altri possono occasionalmente essere positivi, i test commerciale per gli anticorpi è spesso di scarso valore diagnostico. Studi recenti hanno coinvolto autoanticorpi finora sconosciuto antigeni di superficie neuronali (Blaes, Fuhlhuber et al. 2005)
l’Opsoclonus possono anche essere causati da un’infezione virale del tronco encefalico e del cervelletto, così come processi autoimmuni. Un enorme ondata di opsoclono sono comparsi a Chicago, nel 2003 dopo scoppio di una pandemia del virus del Nilo occidentale . Tuttavia, , dopo che lo screening per il cancro è stato negativo, si può pensare a questo tipo di diagnosi.
L’Opsoclono è classicamente attribuito al malfunzionamento delle cellule-pausa del tronco encefalico della linea mediana. Tuttavia, la risonanza magnetica funzionale in due pazienti ha suggerito una aumentata attivazione nei nuclei profondi del cervelletto nell’opsoclono (Helmchen et al, 2003). La regione oculomotor fastigiale ,i neuroni scoppio, i neuroni omnipause e l’anello di retroazione locale sono i generatori del tronco encefalico delle saccade. La regione oculomotor fastigiale è inibita dal verme. Così il problema di fondo fondamentale dell’opsoclono può riguardare una diminuzione di attivazione del lobulo posteriore del Verme.
La Sindrome Opsoclono mioclono, nota anche come atassia Opsoclono-mioclono, è una rara malattia neurologica di cause sconosciute, ma che sembra essere il risultato di un processo autoimmune che coinvolge il sistema nervoso. Si tratta di una condizione estremamente rara. Colpisce 2-3% dei bambini con neuroblastoma.
I sintomi includono: opsoclono, mioclono, atassia cerebellare, afasia, mutismo, letargia, irritabilità o malessere, strabismo, vomito, disturbi del sonno.
Il trattamento si basa sull’ asportazione della neoplasia e somministrazione di immunoglobuline, cortisone e rituximab.
Che cosa è la sindrome opsoclono-mioclono?
La malattia neurologica rara chiamata sindrome opsoclono-mioclono è una malattia autoimmune che attacca il cervello, spesso dopo una infezione virale. Conosciuto anche come “occhi danzanti / piedi danzanti” o sindrome Kinsbourne, questo disturbo può colpire pazienti di qualsiasi età, prima che si manifesta in spasmi agli occhi e muscoli. Alla fine, in piedi o camminare diventa affannoso, come fa a parlare, mangiare o pensare chiaramente.
La sindrome opsoclono-mioclono è segnato da quattro condizioni diverse, che possono apparire quasi istantaneamente. Opsoclono riferisce a contrazioni incontrollabili degli occhi, causando un effetto di danza. Mioclono riguarda spasmi muscolari che vanno e vengono, che interessano tutto il corpo. Anche comune per chi soffre di questa sindrome è atassia, una generale mancanza di coordinazione muscolare che impedisce i movimenti più comuni, dalle passeggiate a mangiare. Infine, encefalopatia ostacola il cervello di pensare chiaramente. Tutto si combinano per creare un paziente che non può controllare il movimento, è letargico, e spesso pieno di rabbia.
Secondo il National Pediatric Mioclono Center, sindrome opsoclono-mioclono segue spesso dopo che il corpo è stato combattendo una crescita tumorale o infetto da varie infezioni virali. I tumori si trovano in circa metà dei pazienti affetti da esso, e le infezioni virali sono stati recentemente trattati in molti del resto. Anche se nel 2011 la causa esatta di questa malattia è sconosciuta, molti ricercatori ritengono che gli anticorpi prodotti dal sistema immunitario del corpo possono inavvertitamente attaccare le cellule del cervello, nel tentativo di liberare il corpo di un tumore invasore o virus.
I medici che curano la sindrome opsoclono-mioclono primo tentativo di determinare se un tumore è presente ordinando una serie di esami radiologici. Se si trova un tumore, che tenteranno di rimuoverlo. Questo spesso elimina la sindrome in childeren, dal momento che il corpo non avverte una presenza invadente. Per gli adulti, tuttavia, la condizione è più probabile continuare anche dopo un tumore è stato rimosso.
Indipendentemente dal fatto che un tumore viene trovato, i medici prescrivono regolarmente un regime di colpi di ormone adrenocorticotropo per trattare la sindrome opsoclono-mioclono. In una serie di iniezioni più di 20 giorni, questo ormone provoca ghiandola surrenale del corpo di produrre più cortisolo, che si traduce in un 80 per cento al 90 per cento tasso di recupero per i bambini, con meno percentuale di recupero per gli adulti.
Altri farmaci possono essere prescritti per spingere questa sindrome in remissione. Immunoglobuline endovenose sono anticorpi umani che possono aumentare la risposta immunitaria del corpo. Chemioterapie e delle terapia steroidea anche sono comunemente consigliato. Sebbene la maggior parte dei pazienti guarisce, molti sono suscettibili di una recidiva quando il corpo sperimenta una nuova malattia.
Molto di più di flutter oculare e opsoclono può essere trovatoqui .
Una rara causa di saccadi che sono troppo veloci per le loro dimensioni sono disturbi oculari in cui è il movimento degli occhilimitato . Una grande saccade può essere programmata centralmente, ma perché l’occhio viene portata breve per restrizione muscolare o rapido affaticamento, una piccola saccade è fatto con la velocità appropriata per un saccade grande. Un indizio è che questi pazienti non fanno mai saccades superi il limite superiore del valore normale per grandi saccades (circa 750 g / sec per registrazioni fatte con una larghezza di banda di 40 Hz)
Cause di patologia (rallentamento) dei saccadici
assunzione di Droghe (molto comune)
Sonnolenza o stanchezza (comune)
Sindromi gangli basali
Corea di Huntington (paresi globale dei saccadici)
Paralisi sopranucleare progressiva (PSP) – (verticale prima)
Morbo di Wilson (estremamente raro)
Sindromi cerebellari
Atrofia olivopontocerebellare
Telangiectasia Atassia
Malattia di Giuseppe (SCA-3)
Debolezza periferica Oculomotoria (paralisi dei nervi cranici, miopatia)
Paralisi del III° e VI° nervo (comune) (link al sesto nervo )
sindrome di Miller Fisher (rara, rallentamento dei saccadici più areflessia e atassia)
Oftalmoplegia Progressiva esterna (PEO) – ![]() vedi film .
vedi film .
Miopatia mitocondriale (estremamente raro)
Disturbi della tiroide (comune, soprattutto verticale)
Sindrome miastenica di Lambert-Eaton (estremamente raro)
miastenia gravis (raro, oscilla)
Malattie della sostanza bianca
Adrenoleucodistrofia – uno Stato membro come la malattia rara.
Oftalmoplegia internucleare (comune nella SM, colpisce di adduzione (in entrata) saccadi principalmente) ![]() film Ino
film Ino
PPRF (lesioni della formazione reticolare parapontina), ad esempio a causa di ms (sclerosi multipla) o ictus cerebrale.
Disturbi vari
Malattia di Niemann-Pick (molto rara)
Oftalmoplegia di Wernicke (estremamente raro)
A seguito di chirurgia cardiaca (globale paresi saccadici)
Malattia di Kennedy (SMA-4, estremamente raro)
Esistono tre tipi di disturbi per la velocità dei movimenti saccadici.
I saccadi possono essere troppo lenti, troppo veloci, o hanno sostanzialmente una velocità o direzione diversa in un occhio rispetto all’altro. Viene diagnosticata un rallentamento saccadici quando velocità media saccadica per un’ampiezza di particolare è inferiore al percentile inferiore quinto normale. La tabella 3 elenca le cause di saccadi lente (da Hain, 1993).
Quando si osserva un rallentamento dei saccadici, si dovrebbe prendere in prima considerazione l’ingestione di farmaci. Gli anticonvulsivanti, sedativi e gli antidepressivi sono generalmente la causa più comune. I movimenti saccadici possono subire un rallentamento fino al 50% quando i pazienti presentano sonnolenza. Se il paziente è sveglio e non prendere un farmaco ad azione centrale, le diagnosi alternative riportate nella tabella 3 debbono essere prese in considerazione .
Sottocategorie dei disturbi saccadici dal loro modello direzionale: paresi globale dei saccadici
In alcuni disturbi, sottigliezze del modello di rallentamento saccadico si permettono di ridurre ulteriormente l’elenco delle possibilità diagnostiche. Si dovrebbe cercare di giudicare se il rallentamento coinvolge tutte le saccadi, o semplicemente le saccadi orizzontali o verticali. Condizioni metaboliche, come l’acquisizione di droga e sonnolenza, possono causare paresi saccadica globale . La Maggior Parte delle condizioni degenerative del sistema nervoso centrale che sono accompagnati da saccadi lente, come le degenerazioni cerebellari ,la corea di Huntington , e le oftalmoplegia esterne croniche progressive possono anche causare un modello simile globale di rallentamento. La sindrome di Miller Fisher, caratterizzata dalla triade oftalmoplegica, atassia ed areflessia è un altro esempio. Allo stesso modo, l’ischemia del tronco cerebrale associata a chirurgia cardiaca possono occasionalmente causare una paresi saccadica globale (Eggers et al, 2008).
![]() Materiale supplementare sul sito DVD : Video di oftalmoplegia esterna progressiva A questo paziente è stata effettuata la diagnosi di sindrome di Kearns-Sayre . Si noti che mentre si muove la testa, gli occhi non ruotano in senso opposto.
Materiale supplementare sul sito DVD : Video di oftalmoplegia esterna progressiva A questo paziente è stata effettuata la diagnosi di sindrome di Kearns-Sayre . Si noti che mentre si muove la testa, gli occhi non ruotano in senso opposto.

Fig. 28
Deficit selettivi nelle saccadi verticali.
D’altra parte, diversi disturbi influenzano saccadi verticali e orizzontali saccadi differenziale. La figura sopra mostra saccadi dello stesso paziente con parkinsonismo. Saccadi verso il basso sono più lenti di saccadi verso l’alto e anche con latenza più lunga. I disordini che colpiscono le saccadi verticali in misura maggiore rispetto alle saccadi orizzontali sono i disturbi del mesencefalo, come la paralisi sopranucleare progressiva (PSP, film e vedi figura sotto), e il coinvolgimento muscolo oculare tipica della malattia della tiroide .
viene riportato che anche la SLA presenta saccadi verticali rallentate (Averbuch Heller et al, 1998 riportarono 2 pazienti con SLA e saccadi verticali lenti. L’autopsia ha mostrato perdita di cellule nel RIMLF e substantia nigra. Okuda, Yamamoto, et al. (1992) riportarono 2 pazienti con saccadi lenti e un paralisi sopranucleare sguardo verticale. L’autopsia in un paziente ha rivelato degenerazione della sostanza nera. Il complesso ALS-Parkinson-demenza è un noto gruppo di pazienti in Guam che combinano le caratteristiche della SLA, e paralisi sopranucleare progressiva .), vedi qui per ulteriori informazioni . Un altro punto utile che può facilitare l’identificazione dei disturbi PSP e affini è la costrizione della gamma .Un esempio di patologia che colpisce le saccadi orizzontali in misura maggiore rispetto alle saccadi verticali è rappresentato dalle lesioni focali quali l’oftalmoplegia internucleare, la paralisi del VI° nervo e nei disturbi dei muscoli oculari laterale e mediale del ponte. Timothy C. Hain
|
|
|
Fig. 29 Saccadi lenti verso il basso in un paziente con PSP . Questi sono stati registrati con una bobina occhio sclerale . |

![]() Materiale supplementare sul sito DVD : Video di saccadi lenti verticali in paziente con PSP (10 mega)
Materiale supplementare sul sito DVD : Video di saccadi lenti verticali in paziente con PSP (10 mega)
Deficit selettivi nelle saccadi orizzontali
Le patologie che possono pregiudicare le saccadi orizzontali in misura maggiore rispetto alle saccadi verticali sono molto rare e comprende i danni del tronco cerebrale associata ad emorragia pontino (sanguinamento) o similari gli effetti non localizzati della chirurgia cardiaca (Vaughn et al, 2008). Un disturbo PSP-simile è stata riportata dopo la riparazione chirurgica dell’aorta ascendente (Mokri et al, 2004). Questa condizione è molto simile PSP, ma sembra in poche settimane o mesi che seguono questo tipo di chirurgia.
|
|

Fig. 30 Saccadi lente verso il basso in un paziente con una PSP Paralisi Sopranucleare Progressiva come la sindrome in seguito a chirurgia aortica (dati inediti di Dr. Hain). Questi sono stati registrati con un sistema di oculografia infrarossi
Deficit che coinvolgono gli occhi particolari o verso l’interno verso l’esterno vs
In oftalmoplegia internucleare, per i movimenti orizzontali, l’occhio di adduzione è rallentato mentre l’occhio sequestro ha tipicamente velocità normale e ha un nistagmo. Questo argomento è trattato in modo molto più dettagliato di seguito sotto il titolo di rallentamento asimmetrico saccadico. Per i movimenti oculari verticali, la FML sembra presentare in primo luogo i segnali sui canali posteriori e anteriori movimenti oculari canali derivati sono risparmiati (Cremer et al, 1999). Le interruzioni di fusione binoculare possono verificarsi in pazienti con INO (Mills et al, 2008).
La Miastenia oculare può determinare debolezza di tutti i muscoli oculari, o essere limitata a singoli muscoli. Così, la distinzione delle saccadi orizzontali-verticali non aiuta nella diagnosi. Piuttosto, la diagnosi è di solito fatta attraverso l’osservazione di fluttuazione dell’ allineamento oculare che varia da un minuto all’altro , la limitazione dei movimenti oculari per la gamma centrale, e la deriva post-saccadica (vedi nella sezione seguente un esempio di deriva post-saccadica ). I miastenici possono anche sviluppare un rallentamento progressivo delle saccadi nel corso del tempo in seguito alla fatica (Spooner e Baloh, 1979). La miastenia oculare è di solito associata con ptosi oculari che di solito vengono colpiti da piccole quantità di edrofonio per via endovenosa (il test del “Tensilon”), ma anche prove positive di Tensilon oculare sono stati riscontrate nella sindrome di Eaton-Lambert (Dell’Osso et al, 1983 ). La sindrome di Eaton-Lambert si presenta comunemente con coinvolgimento oculare (Burns et al, 2003)
La paralisi dei nervi cranici – come la paralisi dei nervi VI° e III ° di solito colpisce un solo occhio e un sottoinsieme dei muscoli oculari.
References
↵ Abadi RV, Gowen E (2004) Characteristics of saccadic intrusions. Vision Res 44:2675–2690. CrossRefMedlineGoogle Scholar
↵ Antoniades CA, Bak TH, Carpenter RH, Hodges JR, Barker RA (2007) Diagnostic potential of saccadometry in progressive supranuclear palsy. Biomark Med 1:487–490. CrossRefMedlineGoogle Scholar
^ Jump up to:ab Bahill, A. Terry; Clark, Michael R.; Stark, Lawrence (1975).“The Main Sequence, A Tool for Studying Human Eye Movements” . Mathematical Biosciences 24 (3-4): 191. doi :10.1016/0025-5564(75)90075-9 .
^ Baloh, Robert W.; Sills, Andrew W.; Kumley, Warren E.;Honrubia, Vicente (1975). “Quantitative measurement of saccade amplitude, duration, and velocity” . Neurology 25(11): 1065. doi : 10.1212/WNL.25.11.1065 .
^ Behrens, Frank; MacKeben, Manfred; Schröder-Preikschat, Wolfgang (2010). “An improved algorithm for automatic detection of saccades in eye movement data and for calculating saccade parameters” . Behavior Research Methods 42 (3): 701. doi : 10.3758/BRM.42.3.701 .
Burns TM and others. Oculobulbar involvement is typical with Lambert-Eaton myasthenic syndrome. Ann Neurol 2003:53:270-273
Cassin, B. and Solomon, S. Dictionary of Eye Terminology. Gainesville, Florida: Triad Publishing Company, 1990[ page needed ]
Cremer PD and others. Ann Neurol 1999:45:529-533.
^ Ebisawa, Y.; Minamitani, H.; Mori, Y.; Takase, M. (1988).“New methods for removing saccades in analysis of smooth pursuit eye movement” . Biological Cybernetics 60 (2): 111.doi : 10.1007/BF00202898 .
Eggers SDZ and others. Selective saccadic palsy after cardiac surgery. Neurology 2008:70:318-320, and Neurology 2008:71:1746-1747 (correspondence)
^ Engbert, Ralf; Kliegl, Reinhold (2003). “Microsaccades uncover the orientation of covert attention”. Vision Research 43(9): 1035–45. doi : 10.1016/S0042-6989(03)00084-1 . PMID12676246 .
^ Jump up to:ab Fischer, B.; Boch, R. (1983). “Saccadic eye movements after extremely short reaction times in the monkey”. Brain Research 260 (1): 21–6. doi : 10.1016/0006-8993(83)90760-6. PMID6402272 .
^ Jump up to:ab Fischer, B.; Ramsperger, E. (1984). “Human express saccades: Extremely short reaction times of goal directed eye movements”. Experimental Brain Research 57 . doi :10.1007/BF00231145 .
Hain TC: Oculomotor Testing: Interpretation. In Handbook of Balance Function Testing (Ed. Jacobson and Newman), Mosby, 1992
^ Hamm, JP; Dyckman, KA; Ethridge, LE; McDowell, JE;Clementz, BA (2010). “Preparatory Activations across a Distributed Cortical Network Determine Production of Express Saccades in Humans” . Journal of Neuroscience 30 (21): 7350–7. doi : 10.1523/JNEUROSCI.0785-10.2010 . PMC3149561 . PMID20505102 .
^ Hopp, J.Johanna; Fuchs, Albert F (2004). “The characteristics and neuronal substrate of saccadic eye movement plasticity”.Progress in Neurobiology 72 (1): 27–53. doi :10.1016/j.pneurobio.2003.12.002 . PMID15019175 .
^ Ibbotson, MR; Crowder, NA; Cloherty, SL; Price, NSC;Mustari, MJ (2008). “Saccadic Modulation of Neural Responses: Possible Roles in Saccadic Suppression, Enhancement, and Time Compression”. Journal of Neuroscience 28 (43): 10952–60. doi : 10.1523/JNEUROSCI.3950-08.2008 . PMID18945903 .
^ Javal, É (1878). “Essai sur la physiologie de la lecture”.Annales d’Oculistique 80 : 61–73.
Knobler RL, Somasundaram M, Schutta HS: Inverse ocular bobbing. Ann Neurol 9:194, 1981
^ Kommerell, G; Olivier, D; Theopold, H (1976). “Adaptive programming of phasic and tonic components in saccadic eye movements. Investigations of patients with abducens palsy”.Investigative ophthalmology 15 (8): 657–60. PMID955831 .
^ Land, MF (1999). “Motion and vision: Why animals move their eyes”. Journal of comparative physiology. A, Sensory, neural, and behavioral physiology 185 (4): 341–52. doi :10.1007/s003590050393 . PMID10555268 .
^ Marple-Horvat, Dilwyn E.; Gilbey, Sean L.; Hollands, Mark Andrew (1996). “A method for automatic identification of saccades from eye movement recordings” . Journal of Neuroscience Methods 67 (2): 191. doi : 10.1016/0165-0270(96)00049-0 .
Mills DA and many others. Break in binocular fusion during head turning in MS patients with INO. Neurology 71, 2008, 458-460
^ Neil R., Carlson, and Donald Heth C. “5.” Psychology: the science of behaviour , fourth Canadian edition. Toronto: Pearson, 2010. 140–141.
^ Pettigrew, JD; Wallman, J; Wildsoet, CF (1990). “Saccadic oscillations facilitate ocular perfusion from the avian pecten”.Nature 343 (6256): 362–3. doi : 10.1038/343362a0 . PMID14756148 .
^ Pierrot-Deseilligny, Charles; Rivaud, Sophie; Gaymard, Bertrand; Muri, Rene; Vermersch, Anne-Isabelle (May 1995).“Cortical Control of Saccades” . Annals of Neurology 37 (5): 557–567. doi : 10.1002/ana.410370504 . PMID7755349 .
^ Jump up to:ab Rommelse, Nanda NJ; Van Der Stigchel, Stefan; Sergeant, Joseph A. (2008). “A review on eye movement studies in childhood and adolescent psychiatry”. Brain and Cognition 68(3): 391–414. doi : 10.1016/j.bandc.2008.08.025 . PMID18835079 .
^ Jump up to:abc “Sensory Reception: Human Vision: Structure and function of the Human Eye” vol. 27, p. 179 Encyclopaedia Britannica, 1987
Thurtell M, Pioro E, Leigh RJ. Abnormal eye movements in Kennedy disease. Neurology 72, 1528-1530, 2009
Troost BT, Daroff RB. (1977) The ocular motor defects in progressive supranuclear palsy. Ann Neurol 2:397-403.
Vaughn et al. Selective saccadic palsy after cardiac surgery. Neurology 2008:71:1746-1747 References [ edit ]
· (correspondence)
Disturbi della precisione saccadici
· Dismetria Overshoot
· Dismetria Undershoot
· Vicino la cecità con superamenti e sottoelencazioni
· Glissadici
· Pulsione
Di questo elenco, superamento dismetria è importante, ma rara. E ‘qualcosa che l’operatore di una macchina ENG deve essere in grado di riconoscere – -Commercial ENG punteggio è quasi inutile. Speriamo che alla fine i produttori di software ITA produrranno un prodotto utile. Gli altri disturbi elencati qui sono o irrilevanti, estremamente raro, o estremamente vulnerabile alle artefatto tecnico.
Tabella 8 elenca i modelli più comuni di inesattezza saccadici che includono dismetria overshoot, undershoot dismetria, glissades e pulsione. Questi disturbi sono causati sia da disturbi oculari e disturbi del sistema nervoso centrale.
Ci sono diverse insidie di essere a conoscenza di quando si considera la diagnosi di dismetria.
Artefatto Blink è il più fastidioso, perché molti soggetti lampeggiano ad ogni saccade, se non diversamente indicato. Artefatto Blink può essere visto facilmente nelle figure 4 e 6 dove ci sono brevi deviazioni nella traccia verticale, della durata di circa 200 msec, accompagnato da deviazioni sincrone in tracce orizzontali. Lampeggia contribuiscono un artefatto tecnico a causa di interazioni con l’EOG e metodi infrarossi per misurare i movimenti oculari. Solo il magnetico sclera tecnica bobina occhio di misurare movimenti oculari è immune a lampeggiare artefatto. Registrazioni EOG è influenzato soprattutto nel piombo verticale, ma nelle registrazioni infrarossi, entrambe le componenti orizzontali e verticali sono interessati. Quando si utilizza registrazioni EOG, è abbastanza comune per la direzione di lampeggio artefatto differiscono tra ciascun occhio, o per lampeggiare artefatto essere forte in un occhio, e assenti nell’altra. Questi problemi sono generalmente connesse ad errori di posizionamento degli elettrodi. Lampeggia sono accompagnati anche da un piccolo movimento dell’occhio (Riggs et al, 1987), e anche possono interagire centrale con saccadi causano overshoot (Hain et al, 1986). Artefatto Blink è meglio evitare di avere una registrazione di piombo verticale disponibile, che permette di ignorare saccadi con lampeggia sovrapposti, e istruendo il paziente per evitare lampeggiare durante i test. Quando un cavo verticale non è disponibile, come nelle figure 2 e 7, è abbastanza difficile essere certi che una saccade di configurazione insolita è veramente aberranti, e si può ripiegare su ispezione visiva diretta del paziente.
Un errore più sottile si riferisce ad errore di calibrazione. Alcuni sistemi elettronistagmografia commerciali calcolano le metriche confrontando lo spostamento saccade effettivo allo spostamento di destinazione. In questa situazione, una taratura errata può causare una dismetria numerico che è un artefatto di errore di calibrazione. Questo errore può essere facilmente individuata controllando le posizioni occhio tracce, come vero dismetria è sempre accompagnato da saccadi correttive.
Un terzo scoglio è la perdita della correzione spettacolo. Le persone che non possono vedere non sono precisi in seguito puntini sullo schermo. Quando si rimuove gli occhiali del paziente per fare il test, è possibile rimuovere anche la loro visione.
|
|
|
Fig. 1: Orizzontale superamento dismetria. Il blu è bersaglio, occhio verde, inferiore rosso, verticale. L’occhio spesso overshoot il bersaglio. Questo paziente aveva una cavernoma di mezzo cerebellare peduncolo di spurgo a sinistra e poi sottoposti a resezione. Non vi è alcun superamento per saccadi verticali. |

Lemij HG, Collewijn H. (1989) Differences in accuracy of human saccades between stationary and jumping targets. Vision Res 29, 1737-1748.
Meienberg O, Zangemeister WH, Rosenberg M, Hoyt WF, Stark L. (1981) Saccadic eye movement strategies in patients with homonymous hemianopia. Ann Neurol 9:537-44.
Rucker JC and others. Neuro-ophthalmology of late-onset Tay-Sachs disease (LOTS). Neurology 2004, 63: 1918-26.
Troost BT, Daroff RB. (1977) The ocular motor defects in progressive supranuclear palsy. Ann Neurol 2:397-403.
Troost BT, Weber RB, Daroff RB. (1972) Hemispheric control of eye movements. 1. Quantitative analysis of refixation saccades in a hemispherectomy patient. Arch Neurol. 27: 441.
Dismetria Overshoot
In dismetria overshoot, la saccade orizzontale iniziale è troppo grande e la saccade correttivo avviene nella direzione opposta allo spostamento di destinazione. Figura 4 mostra dismetria overshoot in un paziente con una lesione cerebellare.
Dismetria Overshoot non è sempre anormale. Nei soggetti normali, transitori overshoot dismetria è comune in saccades rivolte posizione primaria, in saccadi meno di circa 10 gradi in termini di dimensioni, e saccadi fatte ad uno stimolo che appare in una posizione romanzo. I soggetti normali, tuttavia, sarà riaggiustare i loro saccadi per una posizione di destinazione prevedibile e, dopo diversi refixations allo stesso luogo, smettere di produrre superamenti. Dismetria Overshoot è anormale quando è frequente (almeno il 50% del tempo), di dimensioni significative (maggiori di 2 gradi), e quando si verifica in saccades centrifughe superiori a 20 gradi. Mentre i criteri numerici per superamento sono disponibili Weber e Daroff (1971), non ci sentiamo queste
Enduring superamento dismetria è un classico segno di una lesione cerebellare (Selhorst et al, 1976; Ritchie, 1976).
Si può anche verificare nell’occhio sequestro in oftalmoplegia internucleare, nei pazienti con disturbi del campo visivo, e agli occhi del più forte di un abituale fissatore paretico-eye.
· Komiyama A, Toda AH, Johkura K. Edrofonio-indotta oscillazioni macrosaccadic in miastenia grave. Ann Neurol 1999: 45: 522-525
|
|

Fig.4
Undershoot dysmetria– un oculomotor comune “anomalia”
In dismetria undershoot, la saccade iniziale è troppo piccolo e il saccade correttiva prosegue poi verso il bersaglio. Dismetria Undershoot, almeno sia a destra che a sinistra, non porta la stessa connotazione patologica come fa dismetria overshoot come undershoot è comune nei soggetti normali. . I soggetti normali mostreranno circa 1-2 gradi di undershoot per 20 gradi e grandi spostamenti di destinazione (Lemij e Collewijn 1989) undershooting di saccadi è molto comune in contesti clinici – i pazienti con problemi di vista, ad esempio a causa di cataratta o di incapacità a indossare occhiali durante le prove oculomotor, può essere semplicemente indovinando da nuova posizione di destinazione, e può produrre undershoot o sovraelongazione modelli, tipicamente per entrambe le parti. Per questo motivo, sottoelongazioni non è specifico segno di particolare malattia e non eccessivamente molto deve essere fatto di esso sui test ENG.
Costante e significativo (prima saccade <50% del target spostamento) sottoelongazioni è indicativo di un disturbo gangli basali come la malattia o la paralisi sopranucleare progressiva di Parkinson (PSP), e le malattie degenerative del sistema nervoso, come Tay-Sachs (Rucker et al, 2004) .
Undershoot da un lato è soltanto indicativa di un difetto del campo visivo o una lesione cerebellare unilaterale. I dati sopra riportati e belowshows un esempio di saccadi hypometric prodotte dai pazienti con emianopsia sinistra. Nota in cima che vi è una miscela di undershoot e overshoot, e che l’imprecisione è principalmente quello di sinistra. Saccades sono della velocità normale. In fondo, si noti che non vi siano sforamenti, ma ci sono in cerca saccadici a sinistra.
|
|

Fig.5
· Lemij HG, Collewijn H. (1989) Le differenze di accuratezza delle saccadi umani tra gli obiettivi fissi e saltando. Vision Res 29, 1737-1748.
· Meienberg O, Zangemeister WH, Rosenberg M, Hoyt WF, Stark L. (1981) strategie di movimento oculari saccadici nei pazienti con emianopsia omonima. Ann Neurol 9: 537-44.
· Rucker JC e altri. Neuro-oftalmologia di insorgenza tardiva di malattia di Tay-Sachs (LOTTI). Neurology 2004 63: 1918-1926.
· Troost BT, Daroff RB. (1977) I difetti del motore oculari in sopranucleare progressiva paralisi. Ann Neurol 2: 397-403.
· Troost BT, Weber RB, Daroff RB. (1972) il controllo emisferica dei movimenti oculari. 1. L’analisi quantitativa delle saccadi refixation in un paziente emisferectomia. Arch Neurol. 27: 441.
Vicino Cecità:
Sorprendentemente, spesso si dimentica che se le persone non vedenti sono totalmente inesatte, persone quasi cieche possono anche avere problemi nel seguenti piccoli puntini in giro sugli schermi. Inoltre, quando togliamo gli occhiali a questi pazienti, possiamo essere renderli quasi ciechi.
Un esempio interessante di questo è mostrato nella traccia sotto del saccadico, di un paziente affetto da retinite pigmentosa . Le persone con RP eventualmente sviluppare una visione a tunnel. Così, quando il punto viene spostato per il paradigma saccade, una persona con RP deve trovare, come in sostanza, è semplicemente scomparso.
|
|
|
|
|
Fig.6 Saccadi dismetrico nelle persone con retinite pigmentosa (RP), che è una malattia oculare che ostacola la visione periferica. La latenza è allungata, e le saccadi vanno sotto e sopra, a seconda dello stile del paziente . |


{kind=link}
Perché la visione centrale viene risparmiato nei pazienti con RP, lo Smooth Pursuit può essere normale. Questo è illustrato nella pagina che discute i disturbi di monitoraggio.
Pulsione
Il termine “pulsione” viene applicato a saccadi verticali che sono tirati a destra o a sinistra, che richiedono un saccade correttivo orizzontale per fissare il bersaglio. Sia positiva che negativa saccades sono tirati nella stessa direzione orizzontale. Pulsione verso il lato di lesione, o “ipsipulsion”, si verifica dopo infarti nella distribuzione del posteriore inferiore dell’arteria cerebellare (Meyer et al, 1980). PULSION lontano dal lato di lesione, o “contrapulsion”, può verificarsi dopo infarti nella distribuzione dell’arteria cerebellare superiore (Ranalli e Sharpe, 1986). La maggior parte dei laboratori clinici non tentano di registrare pulsione.
· Meyer KT, Baloh RW, Krohel GB, Dow BM. (1980) lateropulsion oculare: un segno di malattia midollare laterale. Arch Ophthal 98, 1614-1616
· Ranalli PJ, Sharpe JA. (1986) Contrapulsion di saccadi e atassia omolaterale: Un disturbo unilaterale del cervelletto rostrale. Ann Neurol 20: 311-316
Glissadici
Il termine “glissade” è un tipo di disturbo della saccade in cui l’occhio non si ferma in un punto fisso, ma piuttosto si muove lentamente oltre il punto.. “Glissades Onward” si verificano quando l’occhio continua a scivolare nella stessa direzione come la parte più veloce della saccade e glissades “indietro” si verificano quando l’occhio vaga in direzione opposta come principale movimento saccadico. Figura 6 illustra indietro glissades in un paziente con miastenia gravis. Glissades si verificano in condizioni in cui il tronco encefalico miscalculates il “Pulse” di attività oculomotoria necessaria per ottenere l’occhio alla nuova posizione o il “passo” di innervazione necessario per tenere l’occhio in atto contro le forze elastiche. Così glissades sono spesso detto di essere a causa di un “disallineamento pulse-step”. I pazienti che hanno rapidi cambiamenti nella funzione oculomotor, come myasthenics oculari sono particolarmente inclini a sviluppare un modello glissadic, perché la quantità di scarica neurale necessaria per ottenere una determinata posizione degli occhi e per tenerlo contro forze antagoniste elastiche è costantemente variabile. Myasthenics possono anche dare prova di una deriva più breve chiamato “faretra” (Yee et al, 1976). Quiver non si verifica nella sindrome di Eaton-Lambert (Dell’Osso et al, 1983). I pazienti con lesioni cerebellari possono produrre glissades perché non sono in grado di regolare il loro rapporto passo impulsi. I pazienti con oftalmoplegia internucleari mostrano glissades poi prolungati negli occhi di adduzione e glissades più breve indietro negli occhi sequestro.
L’insidia principale da considerare quando si cerca di decidere se un paziente ha glissades è l’adeguatezza della stabilizzazione testa. Se la testa è libera di muoversi e lo fa durante una saccade, l’occhio-componente di una saccade testa occhi combinata può assomigliare ad una glissade. Registrazioni infrarossi hanno anche un problema particolare in quanto potrebbero mostrare un artefatto glissade simile relativa a cambiamenti di posizione della palpebra che accompagnano i movimenti saccadici.
· Alpert JN, Coats AC, Perusquia E. (1975) “Saccadic Nystagmus” in cortical cerebellar atrophy.Neurology 25: 276-280.
· Dell’osso LF, Ayyar DR, Daroff RB, Abel LA. Edrophonium test in Eaton-Lambert syndrome: Quantitative oculography. Neurology, September 1983, 33:1157-63.
· Spooner J, Baloh R. Eye movement fatigue in myasthenia gravis. (1979) Neurology, 29:29-33. January.
· Yee RD, Cogan DG, Zee DS, Baloh RW, Honrubia V. (1976) Rapid eye movements in myasthenia gravis. II. Electro-oculographic analysis. Arch Ophthalmol 94: 1465-1472.
References:
Arima K, and others. Two brothers with frontotemporal dementia and parkinsonism with an N279K mutation of the tau gene. Neurology 2000:54:1787-1795
Arvin KL and others. Minocycline markedly protects the neonatal brain against hypoxic-ischemic injury. Ann Neurol 2002:52:54-61
Baker KB, Montgomery EB. Performance on the PD test battery by relatives of patients with progressive supranuclear palsy. Neurology 2001:56:25-30
Bennett and others. Direct genetic evidence for involvement of tau in progressive supranuclear palsy. Neurology 1998:51:982-985
Bensimon, G., A. Ludolph, et al. (2009). “Riluzole treatment, survival and diagnostic criteria in Parkinson plus disorders: the NNIPPS study.” Brain 132 (Pt 1): 156-71.
Biosse V and others. Opthalmologic features of Parkinson’s disease. Neurology 2004:2:177-180
Boroni B and many others. Tau forms in CSF as a reliable biomarker for progressive supranuclear palsy. Neurology 2008:71:1796-1803
Bower JH, Maraganore DM, McDonnell SK, Rocca WA. Incidence of progressive supranuclear palsy and multiple system atrophy in Olmsted County, Minnesota, 1976-1990. Neurology 1997:49:1284-1288
Caparros-Lefebvre D. Possible relation of atypical parkinsonism in the french West INdies with consumption of tropical plants: a case-control study. Lancet 354(9175):281-286, 1999
Chen J and others. Statins induce angiogenecis, neurogenesis and synaptogenesis after stroke. Ann Neurol 2003:53:743-51
Cole DG, Growdon JH. Therapy for progressive supranuclear palsy, past and future. J.Neural Transm 1994:42:283-290
Cordato NJ, Halliday GM, Harding AJ, Hely MA, Morris JGL. Regional brain atrophy in progressive supranuclear palsy and Lewy body disease.
Cox PA, Sacks OW. Cycad neurotoxins, consumption of flying foxes, and ALS-PDC disease in Guam. Neurology 20002 58:956-959
Delacourte A and others. Vulnerable neuronal subsets in Alzheimers and Pick’s disease are distinguished by their tau isoform distribution and phosphorylation. Annals of Neurology 43(2): 193-204, 1998
Di Maria et al. Corticobasal degeneration shares a common genetic background with progressive supranuclear palsy. Ann Neurol 2000:47:374-377
Engel PA (1996). “Treatment of progressive supranuclear palsy with amitriptyline: therapeutic and toxic effects.” J Am Geriatr Soc 44(9): 1072-4.
Foster NL, and others. PET measures of benzodiazepine receptors in progressive supranuclear palsy. Neurology 2000, 54:1768-1773
Fratelli CM, Sonies BC, Chi-Fishman G, Litvin I. Effects of physostigmine on swallowing and oral motor functions in patients with progressive supranuclar palsy: A pilot study. Dysphagia 14(3):165-168, 1999. Comment: this medication did not help.
Goetz CG, Leurgans S, Lang AE and Litvan I (2003). “Progression of gait, speech and swallowing deficits in progressive supranuclear palsy.” Neurology 60(6): 917-22.
Houlden H and others. Corticobasal degeneration and progressive supranuclear palsy share a common tau haplotype. Neurology 2001:56:1702-6.
Glika J, Bogousslavsky J. Presymptomatic hypertension is a major feature in the diagnosis of progressive supranuclear palsy. Arch Neurl 54(9):1104-1108, 1997. Comment: hypertension is so common that this observation, even if true, is of limited significance.
Golbe LI. New survey finds PSP five-fold more common than thought. PSP advocate, 2000 Comment: this study does not really document that PSP is 5 fold more common, this figure is nearly the same as was found in the study of Bower, published in 1997 (TCH). This study actually simply shows that an older study done in 1987 was incorrect.
Golbe LI. PSP research update. (PSP Advocate, 3rd issue, 2004) The 8th international congress on Parkinson’s disease and Movement Disorders, June 2004.
Higgins JJ, Litvan I, Pho LT, Li W, Nee LE. Progresive supranuclear palsy is in linkage disequilibrium with the tau and not the alpha-synuclein gene. Neurology 1998:50:270-273
Higgins JJ, Litvan I, Nee LE, Loveless BS. A lack of the R406W tau mutation in progressive supranuclear palsy and corticobasal degeneration. Neurology 1999:52:404-406
Higgins JL, Adler RL, Loveless JM. Mutational analysis of the tau gene in progressive supranuclear palsy. Neurology 1999:53:1421-1424
Jellinger KA, Bancher C. Neuropathology in: Progressive Supranuclear Palsy: Clinical and Research Approaches, Litvin I, Agid Y (eds) New York:Oxford 1992, 44-88
Kaneko CRS: Effect of ibotenic acid lesions of the omnipause neurons on saccadic eye movements in rhesus macaques. J Neurophysiol 6:2229-2242 (1996)
Karapetyan YE and others. Unique drug screening approach for prion diseases identifies tacrolimus and astemizole as antiprion agents. Proc NAS 2013:110(17): 7044-7049
Katzenschlager R and others. Fourteen-year final report of the randomized PDRG-UK trial comparing three initial treatments in PD. Neurology 2008:71:474-480
Kris J, Gowing G, Julien JP. Efficient three-drug cocktail for disease induced by mutant superoxide dismutates. Ann Neurol 2003:53:429-436
Kuniyoshi, S., DE Riley, DS Zee, SG Reich, C. Whitney and RJ Leigh (2002). “Distinguishing progressive supranuclear palsy from other forms of Parkinson’s disease: evaluation of new signs.” Ann NY Acad Sci 956: 484-6.
Liao K and others. Why do patients with PSP fall ? Evidence for abnormal otolith responses. Neurology 2008; 70:802-9
Litvan I, Sastry N, Sonies B. Characterizing swallowing abnormalities in progressive supranuclear palsy. Neurology 1997:48:1654-1662
Litvan I, Campbell G, Mangone CA, Verny M, McKee A, Chaudhuri K, Jellinger K, Pearce RKB, D’Olhaberriague LD. Which clinical features differentiate progressive supranuclear palsy (Steele-Richardson-Olszewski syndrome) from related disorders ? A clinicopathological study. Brain (1997), 120, 65-74
Litvan, I., M. Phipps, et al. (2001). “Randomized placebo-controlled trial of donepezil in patients with progressive supranuclear palsy.” Neurology 57 (3): 467-73.
Martin and others. Association of single-nucleotide polymorphisms of the Tau gene with late-onset parkinson disease. JAMA 2001:2245-2250
Maurage CA and others. Similar brain tau pathology in DM2/PROMM and DM1/Steinert Disease. Neurology 2005:65:1636-1638
Massey LA and many others. The midbrain to pons ratio. A simple and specific MRI sign of progressive supranuclear palsy. Neurology 2013;80:1856-1861
Mokri B, and others. Syndrome resembling PSP after surgical repair of ascending aorta dissection or aneurysm. Neurology 2004, 62:971-73
Moro DA, Bentivoglio AR. Zolpidem in progressive supranuclar palsy. NEJM 341(7):543-544, 1999. Comment: benzodiazepines are effective in other central ataxias, and might also be effective in PSP
Murrell JR, Koller D and others. Familial multi-system tauopathy with presenile dementia is localized to chromosome 17. Am J. Hum. Gen 61(5):1131-8, 1997
Nagahima Y and others. Movement Disorders 12(5) 691-696, 1997. Comment: This study suggests that PET scans can differentiated CGBD from PSP. Unfortunately, PET scans are so expensive that this information is of minimal clinical value.
Nath U, Ben-Shlomo Y, Thomson RG, Lees AJ and Burn DJ (2003). “Clinical features and natural history of progressive supranuclear palsy: A clinical cohort study.” Neurology 60(6): 910-6.
Oba H and many others. New and reliable MRI diagnosis for progressive supranuclear palsy. Neurology 2005: 64: 2050-2055
Olanow CW, Fahn S, Langston JW, Godbold J. Selegiline and Mortaility in Parkinson’s Disease. Ann Neurol 40,6, Dec 1996.
Otto M and others. Efficacy of flupirtine on cognitive function in patients with CJD: A double-blind study. Neurology 2004:62:714-718
Oskarsson B and others. Stiff eyes in stiff-person syndrome. Neurology 71, 2008, 378-379
Pavior and others. Quantitative MRI measurement of superior cerebellar peduncle in progressive supranuclear palsy. Neurology 2005:64:675-679
Reed LA, Wszolek ZK, Hutton M. Phenotypic correlations in FDTP-17. Neurobiology of Aging 22(2001) 89-107.
Rojo et al. Clinical genetics of familial progressive supranuclear palsy. Brain 122(7) 1233-1245, 1999
Santacruz P, Uttl B, Litvan I, Grafman J. Progresive supranuclear palsy. A survey of the disease course. Neurology 1998:50:1637-1647
Schrag A, Ben-Shlomo Y. Quinn NP. Prevalence of progressive supranuclaear palsy and multiple system atrophy: a cross-sectional study. Lancet 354(9192):1771-1775, 1999.
Schiffmann et al. Prospective study of neurological responses … Ann Neurol 1997, 42:613-621 (Gaucher’s disease).
Schmidt, C., B. Herting, et al. (2009). “Valsalva manoeuvre in patients with different Parkinsonian disorders.” J Neural Transm .
Schneider, A. and E. Mandelkow (2008). “Tau-based treatment strategies in neurodegenerative diseases.” Neurotherapeutics 5 (3): 443-57.
Sergeant N, and others. Neurofibrillary degeneration in progressive supranuclear palsy and corticobasal degeneration: tau pathologies with exclusively “exon 10” isoforms. J. Neurochem 1999;72:1243-49
Seppi K, Schocke MF, Esterhammer R, Kremser C, Brenneis C, Mueller J, Boesch S, Jaschke W, Poewe W and Wenning GK (2003). “Diffusion-weighted imaging discriminates progressive supranuclear palsy from PD, but not from the parkinson variant of multiple system atrophy.” Neurology 60(6): 922-7.
Spillantini MG, Goedert M. Tau protein pathology in neurodegenerative disease. Trends Neurosci 1998:21:428-433
Spillantini MG, Goedert M. Tau and Parkinson Disease. JAMA 2001, 286, 2324-2324
Stamelou, M., A. Reuss, et al. (2008). “Short-term effects of coenzyme Q10 in progressive supranuclear palsy: a randomized, placebo-controlled trial.” Mov Disord 23(7): 942-9.
Steele JC, Richardson JC, Olszewski J. Progressive supranuclear palsy. Arch Neurol 1964:10:333-359.
Suteerawattananon M, MacNeill B, Protas EJ. Supported treadmill training for gait and balance in a patient with progressive supranuclar palsy. Phys Ther 2002:82:485-495
Tanagawa A, Komiyama A, Hasegawa O. Truncal muscle tonus in progressive supranuclear palsy. Journal of Neurol, Neurosurg and Psych, 64(2):190-196, 1998. Comment: This article suggests that the neck is stiff but the trunk is not stiff. This study needs to be repeated and confirmed.
Tsuboi Y, Ahlskog JE and others. Lewy bodies are not increased in progressive supranuclear palsy compared with normal controls. Neurology 2001:57:1675-78
Yamamoto, M. and AH Schapira (2008). “Dopamine agonists in Parkinson’s disease.” Expert Rev Neurother 8(4): 671-7.
Weiner WJ, Minager A, Shulman LM. Pramipexole in progressive supranuclear palsy. Neurology 1999:52:873-874. Comment: this study reported no effect.
Wzzolek Z, Uitti RJ, Hutton M. A mutation in the microtubule-associated protein tau in pallido-nigro-luysian degeneration. Neurology 54, 2028, 2000
Zampieri, C. and RP Di Fabio (2008). “Balance and eye movement training to improve gait in people with progressive supranuclear palsy: quasi-randomized clinical trial.” Phys Ther 88 (12): 1460-73.
Zee DS, Robinson DA, Eng DR: A hypothetical explanation of saccadic Oscillations. Ann Neurol 5:405-414 (1979)
Il movimento saccadico è un movimento oculare rapido volontario. Il suo scopo è di portare sulla fovea l’immagine di oggetti di interesse che compaiono sulla periferia della retina. Esso è caratterizzato da tre parametri che sono: ampiezza, durata e velocità di picco. La durata aumenta in proporzione con l’aumentare dell’ampiezza del movimento, mentre la velocità di picco raggiunge un massimo per ampiezze superiori ai 20°. Altri parametri che descrivono la saccade sono la latenza, la metrica, la traiettoria e la strategia. La latenza è il tempo che separa la comparsa dello stimolo dall’inizio del movimento, dipende sia dall’eccentricità del target sia dal paradigma di stimolazione. La metrica descrive l’accuratezza del movimento, vale a dire il rapporto fra posizione finale dell’occhio e posizione del target: un movimento accurato è definito ortometrico, un movimento che termina oltre il bersaglio si dice ipermetrico mentre uno che si arresta prima del target si dice ipometrico. Le saccadi dismetriche sono seguite da una seconda saccade correttiva con una latenza di circa 100 msec. La traiettoria descrive il modo nel quale la saccade viene eseguita. La strategia indica il modo nel quale una saccade viene programmata e in condizioni fisiologiche è costituita da un unico movimento ortometrico o da due movimenti, il secondo sempre correttivo.
I movimenti saccadici assomigliano alla fase rapida del nistagmo. Se, mentre si guarda un bersaglio visivo, l’immagine di questo si sposta rapidamente sulla fovea, gli occhi conservano la posizione che avevano per circa 200 msec e poi compiono un movimento rapido, saccadico, per riportare l’immagine del bersaglio sulla fovea. Analogamente, si compiono movimenti oculari saccadici, volontari ma spesso non coscienti, quando si esplora lo spazio circostante, per quanto ristretto esso sia. I movimenti saccadici sono molto stereotipati; hanno un andamento standardizzato, caratterizzato da un incremento ed un successivo decremento regolari della velocità di rotazione del globo oculare, che può raggiungere i 900 °/sec (1° = 1 cm a 57 cm di distanza dall’occhio). E’ possibile modificare volontariamente l’ampiezza e la direzione dei movimenti saccadici, ma non la loro velocità. Questa è tale, e la durata del movimento così breve (poche decine di msec; se ne compiono in media 3 al secondo), che le informazioni visive non fanno in tempo a modificarne il decorso una volta che sono iniziati; le eventuali correzioni vengono quindi operate con ulteriori piccoli movimenti saccadici che seguono quello primario.
I movimenti saccadici orizzontali vengono generati a livello della formazione reticolare del ponte, mentre quelli verticali dalla formazione reticolare del mesencefalo. Il cervelletto è anch’esso coinvolto, probabilmente per la modulazione basata sull’esperienza. Anche il collicolo superiore, che riceve fibre retiniche dirette, svolge un ruolo di rilievo nel controllo del movimenti saccadici. L’azione collicolare può essere controllata dai campi oculari frontali (area 8 della corteccia cerebrale), che, a loro volta, possono influenzare i movimenti saccadici anche indipendentemente dal collicolo superiore.
Studio dei movimenti oculari saccadici [van Nechel CH 2000]
Il paziente viene invitato a seguire i punti luminosi che si alternano sul piano orizzontale e poi su quello verticale, in maniera aleatoria, in ampiezza e in tempo di comparsa (fig 11) o clinicamente guardando due oggetti distanziati di 30°-40° di disassiamento, a meno di 50 cm dal punto in cui egli si trova. I saccadici si spostano da un oggetto all’altro, a richiesta dell’esaminatore (fig 12) . Il sistema dei saccadici è un sistema di balistica che permette di dirigere i due occhi su una nuova mira visiva; si tratta quindi di un movimento rapido, il più rapido possibile per la meccanica muscolare e le sue costrizioni viscoelastiche.
Bisogna valutare:
■ la latenza di esecuzione del movimento saccadico, ovvero l’intervallo tra la comparsa del segnale luminoso e la partenza del movimento saccadico; tale valutazione deve essere confrontata in ciascun occhio e in ognuna delle direzioni, a sinistra, a destra, verso l’alto e verso il basso; si possono quindi osservare anche dei ritardi nell’esecuzione dei movimenti, dato che l’anomalia massima viene rilevata nelle negligenze di un emicampo visivo, dove la latenza può essere pari a 1 secondo rispetto a un ritardo abituale di meno di 200 millisecondi;
■ l’analisi delle velocità, che permette di valutare una possibile oftalmoplegia internucleare (pseudoparalisi del muscolo retto interno) o un rallentamento generale dei movimenti saccadici in tutte le direzioni, come si nota nella sindrome di Steele-Richardson-Olszewski;
■ la congruenza dei movimenti dei due occhi contemporaneamente; bisogna guardare i due occhi insieme fissando il proprio sguardo sulla radice del naso del paziente e valutando anche, nel proprio campo visivo periferico, la velocità degli occhi del paziente; si osserva in tal modo anche una certa differenza di fase delle velocità: l’occhio che rallenta è patologico;
■ la precisione del movimento saccadico;
■ il mantenimento sul nuovo punto di fissazione.
Dismetrie
Può trattarsi di ipometrie dove il movimento si arresta troppo presto. Dopo un intervallo compreso tra 50 e 200 millisecondi v’è un nuovo saccadico di rifissazione (a volte anche più di uno). Si parla allora di saccadici frammentati.
Può trattarsi di saccadici ipermetrici; l’occhio supera la mira e la ritrova dopo un secondo saccadico di senso opposto, o anche diversi in oscillazioni. Si distinguono diversi tipi di saccadici ipermetrici:
■ movimenti saccadici ipermetrici a onde quadrate; l’occhio è mantenuto sulla mira per 200 millisecondi; le strutture del sistema nervoso centrale ricalcolano un nuovo saccadico di correzione [1, 48];
■ movimenti saccadici con ritorno slittato, generalmente esponenziale;
■ saccadici dei due occhi come quelli del flutter oculare [86];
■ saccadici constatati unicamente sull’occhio in abduzione e generalmente associati a un’ipometria sull’occhio in adduzione, segno di ipermetria di correzione di un’oftalmoplegia internucleare controlaterale.
Le ipermetrie e le ipometrie possono associarsi, determinando la lateropulsione oculare.
Nel soggetto normale, si può constatare in qualunque momento qualche saccadico ipometrico, poco frequente (1 su 10) e di piccola ampiezza (2° circa).

Il test di controllo del sistema per i movimenti oculari saccadici
Per registrare i movimenti saccadici è necessario disporre di una strumentazione composta da un sistema di evocazione di mire di fissazione (barra a led) e da un sistema di registrazione oculografica dotato di apparecchiatura computerizzata per l’elaborazione delle tracce ottenute. L’esame viene condotto sistemando il paziente seduto davanti ad uno schermo col capo ben fissato. Poiché il sistema di registrazione elettro-oculografico o video si basa sulla valutazione di un segnale bioelettrico (il potenziale corneo-retinico), che è diverso da un soggetto all’altro ed è diverso a seconda dello stato di adattamento alla luce, l’esaminando deve rimanere alcuni minuti nell’ambiente dove si condurrà l’esame per raggiungere una condizione di adattamento stabile. La calibrazione dell’apparecchio di registrazione viene eseguita facendo compiere al soggetto alcuni movimenti saccadici guardando alternatamente led a distanza prefissata. La registrazione vera e propria dei saccadici si effettua facendo eseguire movimenti di fissazione di un led che si accende alternativamente con frequenza ed ampiezza random (non bisogna che il soggetto in esame possa “prevedere” l’accensione del led) sul piano orizzontale in un tempo che varia da 1/2 a 2 sec. e con un angolo di ampiezza che va da 3° a 36 °. Sul segnale di movimento oculare, che si è ottenuto, si potranno eseguire una serie di elaborazioni matematiche o di valutazioni morfologiche che consentiranno di trarre delle considerazioni cliniche. Infatti, l’analisi completa di tutte le possibili variazioni dei parametri quantitativi e qualitativi del movimento saccadico si è rivelata un importante test per la valutazione dell’integrità delle strutture nervose che regolano l’attività oculomotoria riflessa.
Nel soggetto normale, un rallentamento transitono dei parametri del movimento oculare saccadico si osserva in alcune condizioni fisiologiche come lo stato di riposo psico-sensoriale oppure nell’intossicazione da alcool o per effetto di farmaci sedativi che modificano, innalzandola, la soglia della reticolare.
Nella patologia, una riduzione dei valori della velocità dei saccadici compare in tutti i casi in cui risulta alterata l’attività sinaptica a livello delle giunzioni neuro-muscolari come si osserva nella miastenia grave. Un rallentamento dei saccadici risulta inoltre evidente nelle lesioni dell’area nucleare preoculomotoria della reticolare pontina, nelle lesioni del pretetto, delle vie di connessione reticolo-oculomotorie e degli stessi nuclei e dei nervi oculomotori. La sofferenza dei centri sopra- nucleari di controllo dell’oculomotilità si accompagna talvolta a modificazioni prevalentemente morfologiche dei movimenti saccadici che appaiono dismetrici come si osserva nel morbo di Parkinson e nella corea di Huntington. Dismetrici appaiono pure i saccadici (ipo e ipermetrici) in alcune lesioni cerebellari e nelle forme patologiche spino-cerebellari e olivo-ponto-cerebellari. Nelle lesioni corticali unilaterali a livello del lobo frontale, i saccadici appaiono ipometrici quando sono diretti sul lato Opposto a quello del focolaio corticale mentre i saccadici sul piano verticale non presentano alcuna
alterazione. Nella Fig. 33 A-B è riportato un esempio di saccadi normali e alcuni tipi di saccadici patologici. Nelle persone anziane si osservano alterazioni dei movimenti saccadici. L’aumento della latenza e la riduzione del valore massimo della velocità angolare sono probabilmente in rapporto a processi degenerativi di quelle strutture nervose centrali che appartengono al sistema per i movimenti saccadici (Schik G.R. e Coli., 2000).

Baloh RW, Honrubia V Clinical neurophysiology of the vestibular system. Philadelphia : Davis FA, 1979
Larmande P. Oeil et cervelet. Influence du cervelet dans le contrôle de l’oculomotricité. [thèse de doctorat], faculté de médecine de Tours, 1978
Toupet M, De Gramont A, Bacri D, Haguenau M, Pepin B Le flutter-opsoclonie. À propos de trois observations. Rev Neurol 1982 ; 138 : 219-239
Bibliografia
J. Fukushima, T. Akao, S. Kurkin, C.R.S. Kaneko, K. Fukushima. The vestibular-related frontal
cortex and its role in smooth pursuit eye movements and vestibular-pursuit interactions. J Vestib
Res, 2006; 16(1-2): 1-22.
M. Versino, S. Colnaghi. Il saccadico nelle malattie del SNC. Atti congressuali “Aggiornamenti di vestibologia”,3a Edizione; Modena, 10-11 Marzo 2006.
A.F. Fuchs, F.R. Robinson, A Straube. Role of the Caudal Fastigial Nucleus i Saccade Generation. I.Neuronal Discharge Patterns Journal of Neurophysiology, 1993; 70(5): 1723-1740.
S. Ramat, R.J. Leigh, D.S. Zee. L.M. Optican. What clinical disorders tell us about the neural control of saccadic eye movements. Brain, 2007; 130: 10-35.
S. Traccis, D. Zambarbieri. I movimenti di inseguimento lento. Pàtron Editore, Bologna 1994.
S. Traccis, D. Zambarbieri. I movimenti saccadici. Pàtron Editore, Bologna 1992.
R.J. Leigh, D.S. Zee. The neurology of eye movements. 2nd ed., F.A. Davis Company, Philadelphia 1991.
U. Büttner, T. Brandt. Ocular motor disorders of the brain stem. Vol. 1, Nr. 2, August 1992, “Clinical Neurology”, Baillière Tindall, London.
P. Pagnini. Interazioni visuo-vestibolo-oculomotorie. da “Manuale di Audiologia” di M. Del Bo, F. Giaccai,
G. Grisanti, 2a ed., Masson Italia Editori, Milano 1994.
P. Pagnini. Patterns EOG visuo-oculomotori nella patologia tronco-cerebellare. Atti del Simposio
Internazionale “Potenziali evocati uditivi nella patologia del sistema nervoso centrale”, C.R.S. Amplifon,Milano, 26-27 Novembre 1982.
F. Giaccai, P. Pagnini. Patterns elettronistagmografici nelle sindromi vestibolari centrali. da “Les
explorations functionelles objectives en O.R.L.”, relazione al XXIII Congresso della Società Latina di O.R.L.,Bruxelles 1980.TrattatodiVestibologia
References [ edit ]
1. ^ Jump up to:ab Cassin, B. and Solomon, S. Dictionary of Eye Terminology. Gainesville, Florida: Triad Publishing Company, 1990[ page needed ]
2. ^ Javal, É (1878). “Essai sur la physiologie de la lecture”.Annales d’Oculistique 80 : 61–73.
3. ^ Neil R., Carlson, and Donald Heth C. “5.” Psychology: the science of behaviour , fourth Canadian edition. Toronto: Pearson, 2010. 140–141.
4. ^ Jump up to:abc “Sensory Reception: Human Vision: Structure and function of the Human Eye” vol. 27, p. 179 Encyclopaedia Britannica, 1987
5. ^ Pierrot-Deseilligny, Charles; Rivaud, Sophie; Gaymard, Bertrand; Muri, Rene; Vermersch, Anne-Isabelle (May 1995).“Cortical Control of Saccades” . Annals of Neurology 37 (5): 557–567. doi : 10.1002/ana.410370504 . PMID7755349 .
6. ^ Jump up to:ab Fischer, B.; Boch, R. (1983). “Saccadic eye movements after extremely short reaction times in the monkey”. Brain Research 260 (1): 21–6. doi : 10.1016/0006-8993(83)90760-6. PMID6402272 .
7. ^ Jump up to:ab Fischer, B.; Ramsperger, E. (1984). “Human express saccades: Extremely short reaction times of goal directed eye movements”. Experimental Brain Research 57 . doi :10.1007/BF00231145 .
8. ^ Hamm, JP; Dyckman, KA; Ethridge, LE; McDowell, JE;Clementz, BA (2010). “Preparatory Activations across a Distributed Cortical Network Determine Production of Express Saccades in Humans” . Journal of Neuroscience 30 (21): 7350–7. doi : 10.1523/JNEUROSCI.0785-10.2010 . PMC3149561 . PMID20505102 .
9. ^ Jump up to:ab Bahill, A. Terry; Clark, Michael R.; Stark, Lawrence (1975).“The Main Sequence, A Tool for Studying Human Eye Movements” . Mathematical Biosciences 24 (3-4): 191. doi :10.1016/0025-5564(75)90075-9 .
10. ^ Baloh, Robert W.; Sills, Andrew W.; Kumley, Warren E.;Honrubia, Vicente (1975). “Quantitative measurement of saccade amplitude, duration, and velocity” . Neurology 25(11): 1065. doi : 10.1212/WNL.25.11.1065 .
11. ^ Engbert, Ralf; Kliegl, Reinhold (2003). “Microsaccades uncover the orientation of covert attention”. Vision Research 43(9): 1035–45. doi : 10.1016/S0042-6989(03)00084-1 . PMID12676246 .
12. ^ Marple-Horvat, Dilwyn E.; Gilbey, Sean L.; Hollands, Mark Andrew (1996). “A method for automatic identification of saccades from eye movement recordings” . Journal of Neuroscience Methods 67 (2): 191. doi : 10.1016/0165-0270(96)00049-0 .
13. ^ Ebisawa, Y.; Minamitani, H.; Mori, Y.; Takase, M. (1988).“New methods for removing saccades in analysis of smooth pursuit eye movement” . Biological Cybernetics 60 (2): 111.doi : 10.1007/BF00202898 .
14. ^ Behrens, Frank; MacKeben, Manfred; Schröder-Preikschat, Wolfgang (2010). “An improved algorithm for automatic detection of saccades in eye movement data and for calculating saccade parameters” . Behavior Research Methods 42 (3): 701. doi : 10.3758/BRM.42.3.701 .
15. ^ Jump up to:ab Rommelse, Nanda NJ; Van Der Stigchel, Stefan; Sergeant, Joseph A. (2008). “A review on eye movement studies in childhood and adolescent psychiatry”. Brain and Cognition 68(3): 391–414. doi : 10.1016/j.bandc.2008.08.025 . PMID18835079 .
16. ^ Kommerell, G; Olivier, D; Theopold, H (1976). “Adaptive programming of phasic and tonic components in saccadic eye movements. Investigations of patients with abducens palsy”.Investigative ophthalmology 15 (8): 657–60. PMID955831 .
17. ^ Hopp, J.Johanna; Fuchs, Albert F (2004). “The characteristics and neuronal substrate of saccadic eye movement plasticity”.Progress in Neurobiology 72 (1): 27–53. doi :10.1016/j.pneurobio.2003.12.002 . PMID15019175 .
18. ^ Ibbotson, MR; Crowder, NA; Cloherty, SL; Price, NSC;Mustari, MJ (2008). “Saccadic Modulation of Neural Responses: Possible Roles in Saccadic Suppression, Enhancement, and Time Compression”. Journal of Neuroscience 28 (43): 10952–60. doi : 10.1523/JNEUROSCI.3950-08.2008 . PMID18945903 .
19. ^ Land, MF (1999). “Motion and vision: Why animals move their eyes”. Journal of comparative physiology. A, Sensory, neural, and behavioral physiology 185 (4): 341–52. doi :10.1007/s003590050393 . PMID10555268 .
20. ^ Pettigrew, JD; Wallman, J; Wildsoet, CF (1990). “Saccadic oscillations facilitate ocular perfusion from the avian pecten”.Nature 343 (6256): 362–3. doi : 10.1038/343362a0 . PMID14756148 .
Bibliografia[modifica | modifica wikitesto]
1. Findlay, J.M, & Gilchrist, I.D. (2003). Active vision. Oxford University Press: Oxford, UK.
2. Hoffman, J.E, & Subramaniam, B. (1995). The role of visual attention in saccadic eye movement. Perception & Psychophysics.